Dark Septate Endophytic Fungi Associated with Sugarcane Plants Cultivated in São Paulo, Brazil

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Sampling and Isolation of Dark Septate Endophytic Fungi
2.2. Identification of the Isolated Dark Septate Endophytic Fungi
3. Results/Discussion
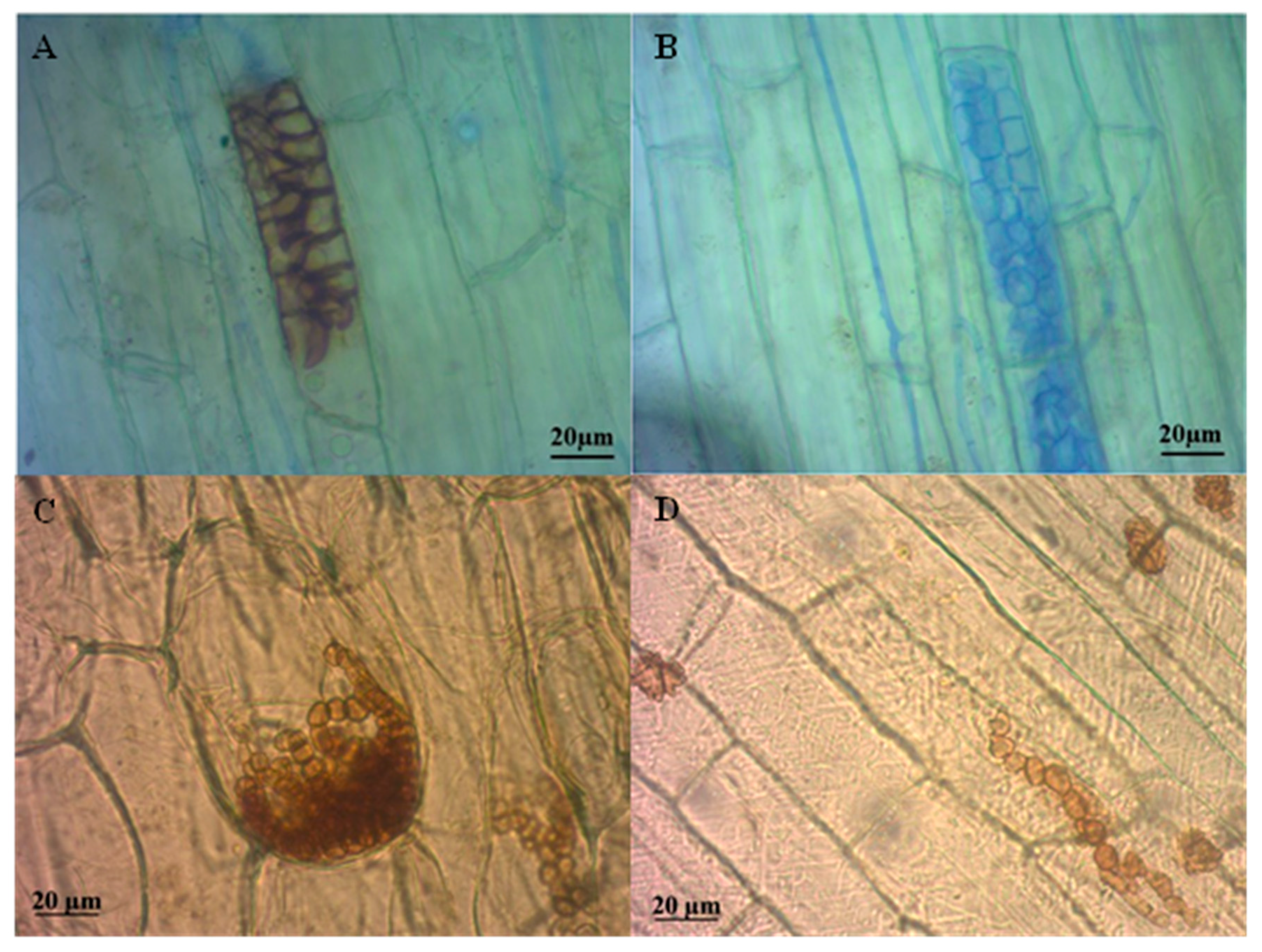
3.1. Isolates of Dark Septate Endophytes
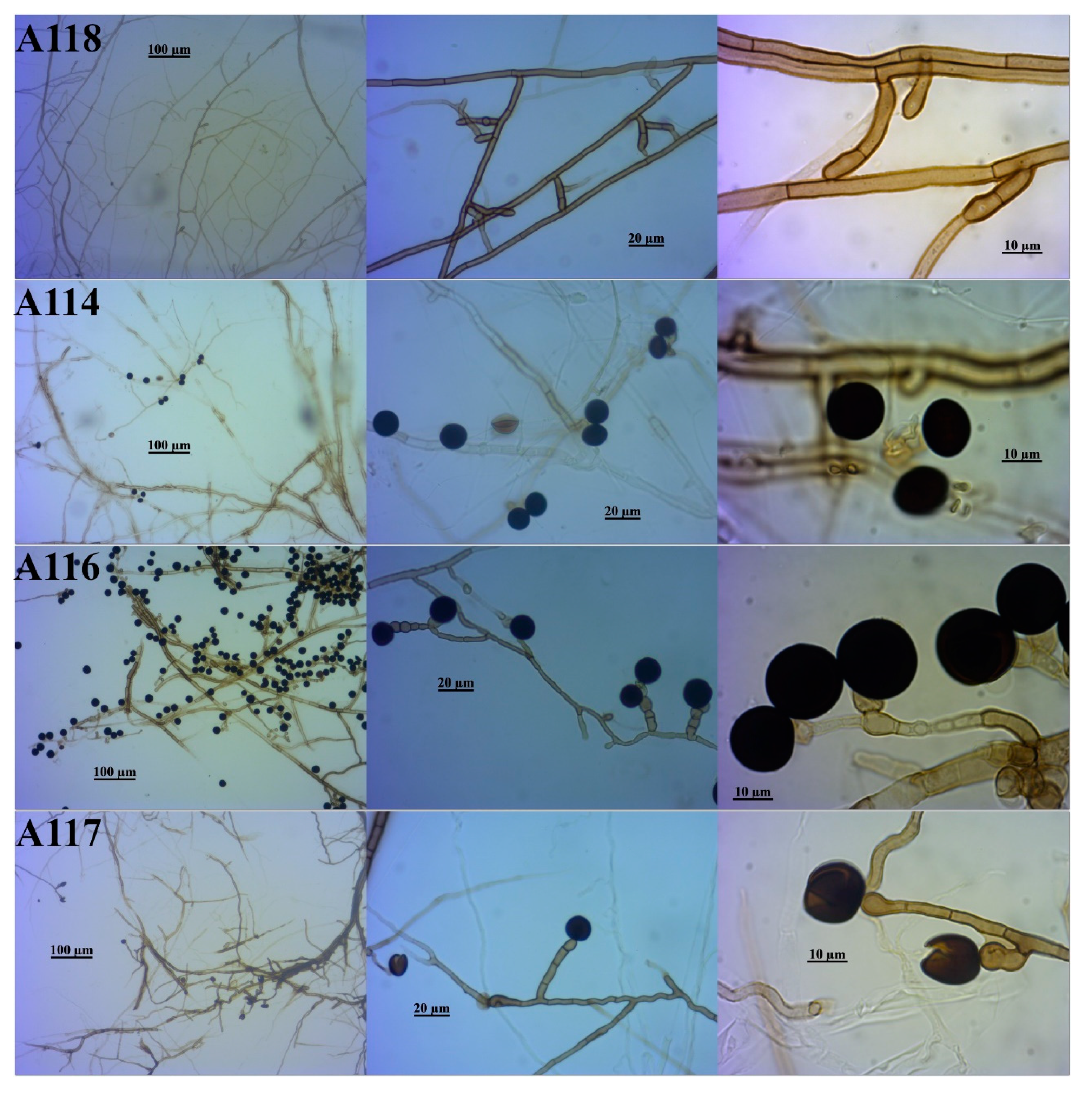
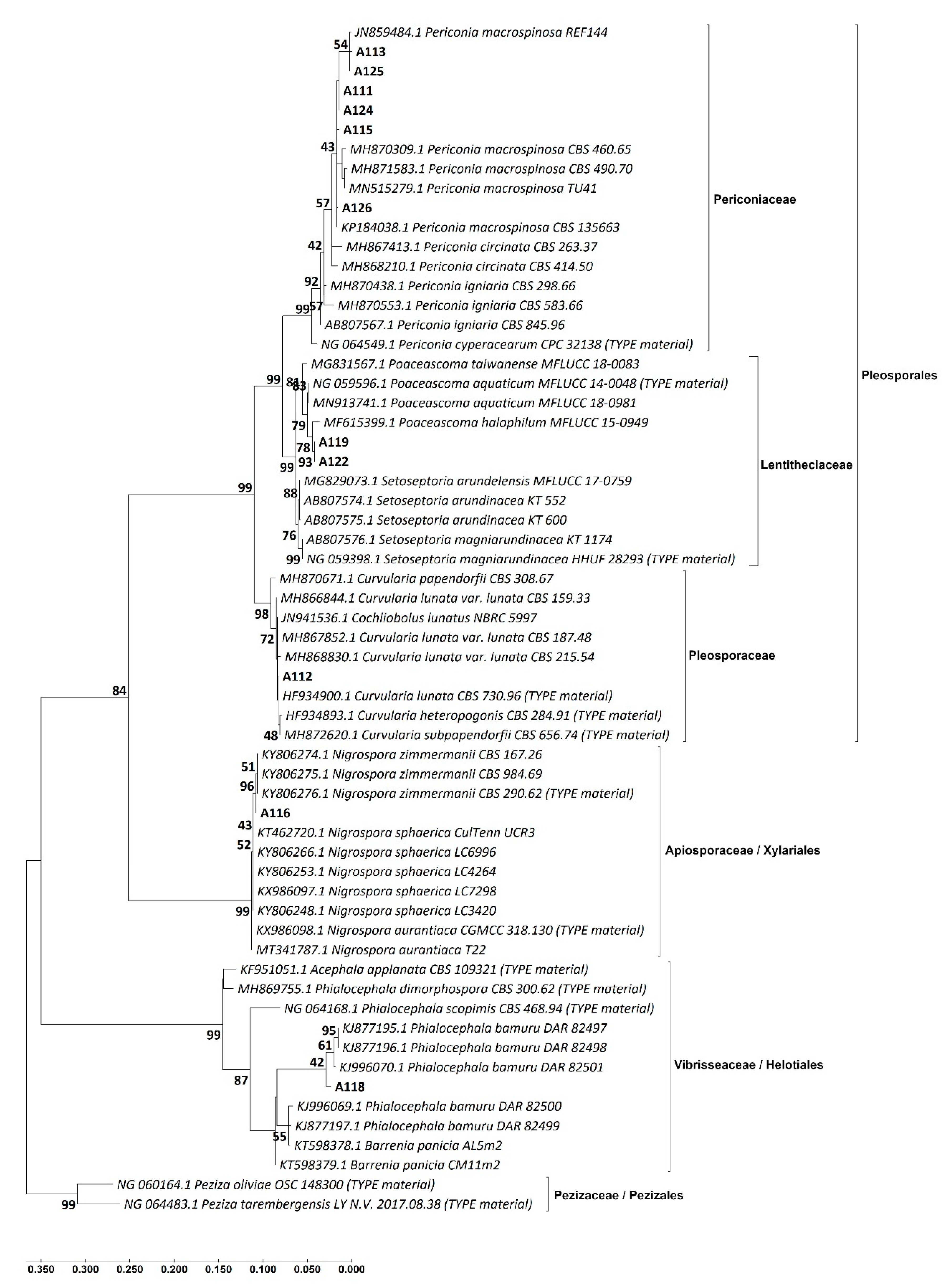
3.2. Identification of the DSE Isolates
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grünig, C.R.; Queloz, V.; Sieber, T.N. Structure of Diversity in Dark Septate Endophytes: From Species to Genes. In Endophytes of Forest Trees: Biology and Applications; Pirttilä, A.M., Frank, A.C., Eds.; Forestry Sciences; Springer: Dordrecht, The Netherlands, 2011; pp. 3–30. ISBN 978-94-007-1599-8. [Google Scholar]
- Ruotsalainen, A.L. Dark Septate Endophytes (DSE) in Boreal and Subarctic Forests. In Endophytes of Forest Trees; Pirttilä, A.M., Frank, A.C., Eds.; Springer International Publishing: Cham, Switzerland, 2018; Volume 86, pp. 105–117. ISBN 978-3-319-89832-2. [Google Scholar]
- Rodriguez, R.J.; White, J.F., Jr.; Arnold, A.E.; Redman, R.S. Fungal endophytes: Diversity and functional roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef]
- Jumpponen, A.; Trappe, J.M. Dark septate endophytes: A review of facultative biotrophic root-colonizing fungi. New Phytol. 1998, 140, 295–310. [Google Scholar] [CrossRef]
- Berthelot, C.; Chalot, M.; Leyval, C.; Blaudez, D. From Darkness to Light: Emergence of the Mysterious Dark Septate Endophytes in Plant Growth Promotion and Stress Alleviation. In Endophytes for a Growing World; Hodkinson, T.R., Doohan, F.M., Saunders, M.J., Murphy, B.R., Eds.; Cambridge University Press: Cambridge, UK, 2019; pp. 143–164. ISBN 978-1-108-60766-7. [Google Scholar]
- Mandyam, K.G.; Jumpponen, A. Mutualism–parasitism paradigm synthesized from results of root-endophyte models. Front. Microbiol. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Knapp, D.G.; Kovács, G.M.; Zajta, E.; Groenewald, J.Z.; Crous, P.W. Dark septate endophytic pleosporalean genera from semiarid areas. Pers. Mol. Phylogeny Evol. Fungi 2015, 35, 87–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonfim, J.A.; Vasconcellos, R.L.F.; Baldesin, L.F.; Sieber, T.N.; Cardoso, E.J.B.N. Dark septate endophytic fungi of native plants along an altitudinal gradient in the Brazilian Atlantic forest. Fungal Ecol. 2016, 20, 202–210. [Google Scholar] [CrossRef]
- Hou, L.; He, X.; Li, X.; Wang, S.; Zhao, L. Species composition and colonization of dark septate endophytes are affected by host plant species and soil depth in the Mu Us sandland, northwest China. Fungal Ecol. 2019, 39, 276–284. [Google Scholar] [CrossRef]
- Knapp, D.G.; Pintye, A.; Kovács, G.M. The Dark Side Is Not Fastidious—Dark Septate Endophytic Fungi of Native and Invasive Plants of Semiarid Sandy Areas. PLoS ONE 2012, 7, e32570. [Google Scholar] [CrossRef] [Green Version]
- Mandyam, K.; Fox, C.; Jumpponen, A. Septate endophyte colonization and host responses of grasses and forbs native to a tallgrass prairie. Mycorrhiza 2012, 22, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Mandyam, K.; Loughin, T.; Jumpponen, A. Isolation and morphological and metabolic characterization of common endophytes in annually burned tallgrass prairie. Mycologia 2010, 102, 813–821. [Google Scholar] [CrossRef] [Green Version]
- Vergara, C.; Araujo, K.E.C.; Urquiaga, S.; Schultz, N.; Balieiro, F.d.C.; Medeiros, P.S.; Santos, L.A.; Xavier, G.R.; Zilli, J.E. Dark Septate Endophytic Fungi Help Tomato to Acquire Nutrients from Ground Plant Material. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Li, T.; Shen, M.; Yang, Z.L.; Zhao, Z.-W. Evidence for a Dark Septate Endophyte (Exophiala Pisciphila, H93) Enhancing Phosphorus Absorption by Maize Seedlings. Plant Soil 2020, 452, 249–266. [Google Scholar] [CrossRef]
- Santos, S.G.; Silva, P.R.A.; Garcia, A.C.; Zilli, J.É.; Berbara, R.L.L. Dark septate endophyte decreases stress on rice plants. Braz. J. Microbiol. 2017, 48, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; He, X.-L.; Zhou, Y.; Hou, Y.-T.; Zuo, Y.-L. Effects of Dark Septate Endophytes on the Performance of Hedysarum scoparium Under Water Deficit Stress. Front. Plant. Sci. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Liu, M.J.; Zhang, X.T.; Zhang, H.B.; Sha, T.; Zhao, Z.W. Improved tolerance of maize (Zea mays L.) to heavy metals by colonization of a dark septate endophyte (DSE) Exophiala pisciphila. Sci. Total Environ. 2011, 409, 1069–1074. [Google Scholar] [CrossRef] [PubMed]
- Likar, M.; Regvar, M. Isolates of dark septate endophytes reduce metal uptake and improve physiology of Salix caprea L. Plant Soil 2013, 370, 593–604. [Google Scholar] [CrossRef]
- Gonzalez Mateu, M.; Baldwin, A.H.; Maul, J.E.; Yarwood, S.A. Dark septate endophyte improves salt tolerance of native and invasive lineages of Phragmites australis. ISME J. 2020, 14, 1943–1954. [Google Scholar] [CrossRef]
- Narisawa, K.; Usuki, F.; Hashiba, T. Control of Verticillium Yellows in Chinese Cabbage by the Dark Septate Endophytic Fungus LtVB3. Phytopathology 2004, 94, 412–418. [Google Scholar] [CrossRef] [Green Version]
- Deng, X.; Song, X.; Halifu, S.; Yu, W.; Song, R. Effects of Dark Septate Endophytes Strain A024 on Damping-off Biocontrol, Plant Growth and the Rhizosphere Soil Enviroment of Pinus sylvestris var. mongolica Annual Seedlings. Plants 2020, 9, 913. [Google Scholar] [CrossRef]
- Harsonowati, W.; Marian, M.; Surono; Narisawa, K. The Effectiveness of a Dark Septate Endophytic Fungus, Cladophialophora chaetospira SK51, to Mitigate Strawberry Fusarium Wilt Disease and With Growth Promotion Activities. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Mandyam, K.G.; Roe, J.; Jumpponen, A. Arabidopsis thaliana model system reveals a continuum of responses to root endophyte colonization. Fungal Biol. 2013, 117, 250–260. [Google Scholar] [CrossRef] [Green Version]
- Rothen, C.; Miranda, V.; Aranda-Rickert, A.; Fracchia, S.; Rodríguez, M.A. Characterization of dark septate endophyte fungi associated with cultivated soybean at two growth stages. Appl. Soil Ecol. 2017, 120, 62–69. [Google Scholar] [CrossRef]
- Mayerhofer, M.S.; Kernaghan, G.; Harper, K.A. The effects of fungal root endophytes on plant growth: A meta-analysis. Mycorrhiza 2013, 23, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Berthelot, C.; Blaudez, D.; Leyval, C. Differential growth promotion of poplar and birch inoculated with three dark septate endophytes in two trace element-contaminated soils. Int. J. Phytoremediat. 2017, 19, 1118–1125. [Google Scholar] [CrossRef] [PubMed]
- Hulse, J.D. First Report of Dark Septate Endophytes imaged in Cucurbita maxima grown in the Eastern United States. Acta Sci. Agric. 2018, 2, 61–64. [Google Scholar]
- Vergara, C.; Araujo, K.E.C.; Urquiaga, S.; Santa-Catarina, C.; Schultz, N.; Silva Araújo, E.; Carvalho Balieiro, F.; Xavier, G.R.; Zilli, J.É. Dark Septate Endophytic Fungi Increase Green Manure-15N Recovery Efficiency, N Contents, and Micronutrients in Rice Grains. Front. Plant. Sci. 2018, 9. [Google Scholar] [CrossRef]
- Rudorff, B.F.T.; Aguiar, D.A.; Silva, W.F.; Sugawara, L.M.; Adami, M.; Moreira, M.A. Studies on the Rapid Expansion of Sugarcane for Ethanol Production in São Paulo State (Brazil) Using Landsat Data. Remote Sens. 2010, 2, 1057–1076. [Google Scholar] [CrossRef] [Green Version]
- Romão-Dumaresq, A.S.; Dourado, M.N.; de Lima Fávaro, L.C.; Mendes, R.; Ferreira, A.; Araújo, W.L. Diversity of Cultivated Fungi Associated with Conventional and Transgenic Sugarcane and the Interaction between Endophytic Trichoderma virens and the Host Plant. PLoS ONE 2016, 11, e0158974. [Google Scholar] [CrossRef]
- Souza, R.S.C.; Okura, V.K.; Armanhi, J.S.L.; Jorrín, B.; Lozano, N.; Silva, M.J.; González-Guerrero, M.; Araújo, L.M.; Verza, N.C.; Bagheri, H.C.; et al. Unlocking the bacterial and fungal communities assemblages of sugarcane microbiome. Sci. Rep. 2016, 6, 28774. [Google Scholar] [CrossRef] [Green Version]
- Claassens, A.; Nock, C.J.; Rose, M.T.; Zwieten, L.V.; Rose, T.J. Colonisation dynamics of arbuscular mycorrhizal fungi and dark septate endophytes in the sugarcane crop cycle. Rhizosphere 2018, 7, 18–26. [Google Scholar] [CrossRef]
- Fors, R.O.; Júnior, O.J.S.; Carneiro, M.A.C.; Berbara, R.L.L. Selection of arbuscular mycorrhizal fungi for sugarcane in four soils with the presence of dark septate endophytes. Acta Sci. Agron. 2020, 42, e42477. [Google Scholar] [CrossRef] [Green Version]
- Santos, H.G.; Jacomine, P.K.T.; Anjos, L.H.C.; Oliveira, V.A.; Lumbreras, J.F.; Coelho, M.R.; Almeida, J.A.; Araujo Filho, J.C.; Oliveira, J.B.; Cunha, T.J.F. Sistema Brasileiro de Classificação de Solos; Embrapa: Brasília, Brazil, 2018; ISBN 978-85-7035-817-2. [Google Scholar]
- Ribeiro, K.G.; Pereira, G.M.D.; Mosqueira, C.A.; Baraúna, A.C.; Vital, M.J.S.; Silva, K.; Zilli, J.É. Isolamento, armazenamento e determinação da colonização por fungos “dark septate” a partir de plantas de arroz. Rev. Agrombiente Online 2011, 5, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Silvani, V.A.; Fracchia, S.; Fernández, L.; Pérgola, M.; Godeas, A. A simple method to obtain endophytic microorganisms from field-collected roots. Soil Biol. Biochem. 2008, 40, 1259–1263. [Google Scholar] [CrossRef]
- Novais, C.B.; Borges, W.L.; Silva, G.A.; Saggin-Júnior, O.J. Técnicas Básicas em Micorrizas Arbusculares; UFLA: Lavras, Brazil, 2017; ISBN 978-85-8127-062-3. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal lRNA genes for phylogenetics. In PCR Protocols; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Rehner, S.A.; Samuels, G.J. Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences. Mycol. Res. 1994, 98, 625–634. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carbone, I.; Kohn, L.M. A Method for Designing Primer Sets for Speciation Studies in Filamentous Ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Groenewald, J.Z.; Nakashima, C.; Nishikawa, J.; Shin, H.-D.; Park, J.-H.; Jama, A.N.; Groenewald, M.; Braun, U.; Crous, P.W. Species concepts in Cercospora: Spotting the weeds among the roses. Stud. Mycol. 2013, 75, 115–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- European and Mediterranean Plant Protection Organization (EPPO) Q-Bank. Available online: https://qbank.eppo.int/fungi/ (accessed on 13 February 2019).
- Morgulis, A.; Coulouris, G.; Raytselis, Y.; Madden, T.L.; Agarwala, R.; Schäffer, A.A. Database indexing for production MegaBLAST searches. Bioinformatics 2008, 24, 1757–1764. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. J. Comput. Mol. Cell Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; ISBN 9780195135855. [Google Scholar]
- MycoBank Database. Available online: https://www.mycobank.org/ (accessed on 17 June 2020).
- Index Fungorum. Available online: http://www.indexfungorum.org/Names/Names.asp (accessed on 17 June 2020).
- Ali-Shtayeh, M.S.; Jamous, R.M.; Yaghmour, R.M.-R. Mycology Manual; An-Najah National University: Nablus, Palestine, 1998. [Google Scholar]
- Lin, W.; Wu, L.; Lin, S.; Zhang, A.; Zhou, M.; Lin, R.; Wang, H.; Chen, J.; Zhang, Z.; Lin, R. Metaproteomic analysis of ratoon sugarcane rhizospheric soil. BMC Microbiol. 2013, 13, 135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; He, C.; He, X.; Su, F.; Hou, L.; Ren, Y.; Hou, Y. Dark septate endophytes improve the growth of host and non-host plants under drought stress through altered root development. Plant Soil 2019, 439, 259–272. [Google Scholar] [CrossRef]
- Yuan, Z.-L.; Zhang, C.-L.; Lin, F.-C.; Kubicek, C.P. Identity, Diversity, and Molecular Phylogeny of the Endophytic Mycobiota in the Roots of Rare Wild Rice (Oryza granulate) from a Nature Reserve in Yunnan, China. Appl. Environ. Microbiol. 2010, 76, 1642–1652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loro, M.; Valero-Jiménez, C.A.; Nozawa, S.; Márquez, L.M. Diversity and composition of fungal endophytes in semiarid Northwest Venezuela. J. Arid Environ. 2012, 85, 46–55. [Google Scholar] [CrossRef]
- Ellis, M.B. Periconia macrospinosa. Descriptions of Fungi and Bacteria. IMI Descr. Fungi Bact. 1968, 17, 168. [Google Scholar]
- Herrera, J.; Poudel, R.; Bokati, D. Assessment of root-associated fungal communities colonizing two species of tropical grasses reveals incongruence to fungal communities of North American native grasses. Fungal Ecol. 2013, 6, 65–69. [Google Scholar] [CrossRef]
- Lima, A.; Furtado, M. Espécies do género Curvularia (fungos anamórficos: Hyphomycetes) na ilha de Santiago, Cabo Verde. Port. Acta Biol. 2007, 22, 145–156. [Google Scholar]
- Manamgoda, D.S.; Cai, L.; McKenzie, E.H.C.; Crous, P.W.; Madrid, H.; Chukeatirote, E.; Shivas, R.G.; Tan, Y.P.; Hyde, K.D. A phylogenetic and taxonomic re-evaluation of the Bipolaris—Cochliobolus—Curvularia Complex. Fungal Divers. 2012, 56, 131–144. [Google Scholar] [CrossRef]
- Liang, Y.; Ran, S.-F.; Bhat, J.; Hyde, K.D.; Wang, Y.; Zhao, D.-G. Curvularia microspora sp. nov. associated with leaf diseases of Hippeastrum striatum in China. MycoKeys 2018, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Bengyella, L.; Iftikhar, S.; Nawaz, K.; Fonmboh, D.J.; Yekwa, E.L.; Jones, R.C.; Njanu, Y.M.T.; Roy, P. Biotechnological application of endophytic filamentous Bipolaris and Curvularia: A review on bioeconomy impact. World J. Microbiol. Biotechnol. 2019, 35, 69. [Google Scholar] [CrossRef]
- Avinash, K.S.; Ashwini, H.S.; Babu, H.N.R.; Krishnamurthy, Y.L. Antimicrobial Potential of Crude Extract of Curvularia lunata, an Endophytic Fungi Isolated from Cymbopogon caesius. J. Mycol. 2015, 2015, 1–4. [Google Scholar] [CrossRef]
- Ricaud, C.; Egan, B.T. (Eds.) Diseases of Sugarcane: Major Diseases; Elsevier: Amsterdam, The Netherlands, 1989; ISBN 0-444-42797-X. [Google Scholar]
- Mehnaz, S. Microbes—Friends and foes of sugarcane: Microbes and sugarcane. J. Basic Microbiol. 2013, 53, 954–971. [Google Scholar] [CrossRef] [PubMed]
- Raza, M.; Zhang, Z.-F.; Hyde, K.D.; Diao, Y.-Z.; Cai, L. Culturable plant pathogenic fungi associated with sugarcane in southern China. Fungal Divers. 2019, 99, 1–104. [Google Scholar] [CrossRef]
- Tanney, J.B.; Douglas, B.; Seifert, K.A. Sexual and asexual states of some endophytic Phialocephala species of Picea. Mycologia 2016, 108, 255–280. [Google Scholar] [CrossRef] [Green Version]
- Wong, P.T.W.; Dong, C.; Martin, P.M.; Sharp, P.J. Fairway patch—A serious emerging disease of couch (syn. bermudagrass) Cynodon dactylon and kikuyu (Pennisetum clandestinum) turf in Australia caused by Phialocephala bamuru P.T.W. Wong & C. Dong sp. nov. Australas. Plant. Pathol. 2015, 44, 545–555. [Google Scholar] [CrossRef]
- Walsh, E.; Luo, J.; Naik, A.; Preteroti, T.; Zhang, N. Barrenia, a new genus associated with roots of switchgrass and pine in the oligotrophic pine barrens. Fungal Biol. 2015, 119, 1216–1225. [Google Scholar] [CrossRef] [Green Version]
- Tanney, J.B.; Seifert, K.A. Mollisiaceae: An overlooked lineage of diverse endophytes. Stud. Mycol. 2020. [Google Scholar] [CrossRef]
- Wang, M.; Liu, F.; Crous, P.W.; Cai, L. Phylogenetic reassessment of Nigrospora: Ubiquitous endophytes, plant and human pathogens. Persoonia 2017, 39, 118–142. [Google Scholar] [CrossRef]
- Hao, Y.; Aluthmuhandiram, J.V.S.; Chethana, K.W.T.; Manawasinghe, I.S.; Li, X.; Liu, M.; Hyde, K.D.; Phillips, A.J.L.; Zhang, W. Nigrospora Species Associated with Various Hosts from Shandong Peninsula, China. Mycobiology 2020, 48, 169–183. [Google Scholar] [CrossRef]
- Zhao, J.H.; Zhang, Y.L.; Wang, L.W.; Wang, J.Y.; Zhang, C.L. Bioactive secondary metabolites from Nigrospora sp. LLGLM003, an endophytic fungus of the medicinal plant Moringa oleifera Lam. World J. Microbiol. Biotechnol. 2012, 28, 2107–2112. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Cai, X.; Pang, Q.; Zhou, M.; Chen, Y.; Zhang, W.; Bian, Q. First Report of Leaf Blight on Mentha canadensis Caused by Nigrospora sphaerica in China. Plant. Dis. 2020. [Google Scholar] [CrossRef]
- Chen, X.; Wang, N.; Yang, M.-F.; Li, H.-X. First Report of Nigrospora Leaf Spot Caused by Nigrospora oryzae on Watermelon in China. Plant. Dis. 2018, 103, 1019. [Google Scholar] [CrossRef]
- Cui, Y.P.; Wu, B.; Peng, A.T.; Li, Z.L.; Lin, J.F.; Song, X.B. First Report of Nigrospora Leaf Blight on Sugarcane Caused by Nigrospora sphaerica in China. Plant. Dis. 2017, 102, 824. [Google Scholar] [CrossRef]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W.; Consortium, F.B. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samson, R.A.; Visagie, C.M.; Houbraken, J.; Hong, S.-B.; Hubka, V.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Susca, A.; Tanney, J.B.; et al. Phylogeny, identification and nomenclature of the genus Aspergillus. Stud. Mycol. 2014, 78, 141–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raja, H.A.; Miller, A.N.; Pearce, C.J.; Oberlies, N.H. Fungal Identification Using Molecular Tools: A Primer for the Natural Products Research Community. J. Nat. Prod. 2017, 80, 756–770. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.-B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baum, C.; Hrynkiewicz, K.; Szymańska, S.; Vitow, N.; Hoeber, S.; Fransson, P.M.A.; Weih, M. Mixture of Salix Genotypes Promotes Root Colonization with Dark Septate Endophytes and Changes P Cycling in the Mycorrhizosphere. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Shrestha, P.; Szaro, T.M.; Bruns, T.D.; Taylor, J.W. Systematic Search for Cultivatable Fungi That Best Deconstruct Cell Walls of Miscanthus and Sugarcane in the Field. Appl. Environ. Microbiol. 2011, 77, 5490–5504. [Google Scholar] [CrossRef] [Green Version]
- Surono; Narisawa, K. The dark septate endophytic fungus Phialocephala fortinii is a potential decomposer of soil organic compounds and a promoter of Asparagus officinalis growth. Fungal Ecol. 2017, 28, 1–10. [Google Scholar] [CrossRef]
- Martins, T.D.; Menten, J.O.M.; Sanguino, Á. Fungos associados às sementes (Cariopses) de cana-de-açúcar: Métodos para detecção, incidência e relação entre incidência fúngica e ambiente de produção das sementes. Summa Phytopathol. 2009, 35, 173–178. [Google Scholar] [CrossRef]
- Hardoim, P.R.; Hardoim, C.C.P.; van Overbeek, L.S.; van Elsas, J.D. Dynamics of Seed-Borne Rice Endophytes on Early Plant Growth Stages. PLoS ONE 2012, 7, e30438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardoim, P.R.; Overbeek, L.S.; Elsas, J.D. Properties of bacterial endophytes and their proposed role in plant growth. Trends Microbiol. 2008, 16, 463–471. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Municipality | pH | OM | P-resin | K | Ca | Mg | H + Al | Al | CTC | Sampled Point | Sugarcane Variety | Plantation Age |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| g dm−3 | mg dm−3 | mmolc dm−3 | ||||||||||
| João Ramalho | 4.66 | 10.4 | 5.2 | 0.98 | 9.60 | 4.80 | 16.8 | 0.6 | 32.2 | 1 | RB867515 | 1st ratoon |
| 2 | RB966928 | 1st ratoon | ||||||||||
| 4 | RB92579 | 1st ratoon | ||||||||||
| Paraguaçu Paulista | 5.43 | 5.0 | 10.5 | 1.30 | 15.17 | 5.17 | 14.0 | 0.5 | 35.5 | 3 | RB867515 | 1st ratoon |
| Paraguaçu Paulista | 5.45 | 10.2 | 16.4 | 2.37 | 17.00 | 6.18 | 14.8 | 0.0 | 40.4 | 11 | RB92579 | 1st ratoon |
| Quatá | 5.58 | 6.1 | 7.0 | 0.96 | 14.31 | 5.92 | 14.5 | 0.9 | 35.8 | 6 | CE2 * | Plant-cane |
| 9 | CE4 * | Plant-cane | ||||||||||
| 12 | CE1 * | Plant-cane | ||||||||||
| 16 | CE3 * | Plant-cane | ||||||||||
| Quatá | 5.70 | 5.0 | 3.0 | 0.53 | 10.67 | 4.67 | 11.0 | 0.0 | 27.3 | 8 | RB867515 | 1st ratoon |
| 10 | RB92579 | 1st ratoon | ||||||||||
| 13 | RB966928 | 1st ratoon | ||||||||||
| Quatá | 6.08 | 12.9 | 15.9 | 1.42 | 17.35 | 5.71 | 12.1 | 0.0 | 36.6 | 15 | RB92579 | 1st ratoon |
| Quatá | 6.20 | 10.5 | 22.2 | 2.63 | 18.50 | 7.33 | 10.8 | 0.0 | 39.3 | 5 | RB867515 | Plant-cane |
| Quatá | 6.20 | 17.0 | 43.5 | 3.55 | 19.25 | 6.75 | 13.2 | 0.0 | 42.7 | 7 | RB966928 | 1st ratoon |
| Quatá | 6.21 | 5.3 | 20.6 | 1.54 | 18.00 | 5.93 | 11.2 | 0.0 | 36.8 | 14 | RB966928 | 1st ratoon |
| Sugarcane Variety | Plantation Age | Sampled Point | Isolated Colonies | ||
|---|---|---|---|---|---|
| Total | Dark | Code Assigned * | |||
| RB867515 | 1st ratoon | 1 | 3 | 2 | A111, A112 |
| 1st ratoon | 3 | 4 | 0 | - | |
| Plant-cane | 5 | 2 | 0 | - | |
| 1st ratoon | 8 | 6 | 3 | A113–A115 | |
| RB92579 | 1st ratoon | 4 | 0 | 0 | - |
| 1st ratoon | 10 | 0 | 0 | - | |
| 1st ratoon | 11 | 3 | 1 | A116 | |
| 1st ratoon | 15 | 2 | 1 | A117 | |
| RB966928 | 1st ratoon | 2 | 12 | 0 | - |
| 1st ratoon | 7 | 8 | 4 | A118–A121 | |
| 1st ratoon | 13 | 2 | 0 | - | |
| 1st ratoon | 14 | 4 | 2 | A122, A123 | |
| CE1 | Plant-cane | 12 | 2 | 0 | - |
| CE2 | Plant-cane | 6 | 7 | 0 | - |
| CE3 | Plant-cane | 16 | 5 | 3 | A124–A126 |
| CE4 | Plant-cane | 9 | 3 | 0 | - |
| Isolate Code * | Identification Based on Morphological Similarity | Closest Identification Based on BLAST and ML Phylogenetic Analysis of | Identification Consensus of This Study | |||
|---|---|---|---|---|---|---|
| ITS Sequences | LSU Sequences | Genus/Species | Family | Order | ||
| Class Dothideomycetes | ||||||
| A111 | Periconia macrospinosa | Periconia macrospinosa | Periconia af. macrospinosa | Periconia macrospinosa | Periconiaceae | Pleosporales |
| A113 | Periconia macrospinosa | Periconia macrospinosa | Periconia af. macrospinosa | Periconia macrospinosa | ||
| A115 | Periconia macrospinosa | Periconia macrospinosa | Periconia af. macrospinosa | Periconia macrospinosa | ||
| A124 | Periconia macrospinosa | Periconia macrospinosa | Periconia af. macrospinosa | Periconia macrospinosa | ||
| A125 | Periconia macrospinosa | Periconia macrospinosa | Periconia af. macrospinosa | Periconia macrospinosa | ||
| A126 | Periconia macrospinosa | Periconia macrospinosa | Periconia af. macrospinosa | Periconia macrospinosa | ||
| A112 | Curvularia sp. | Curvularia lunata | Curvularia af. lunata | Curvularia lunata | Pleosporaceae | |
| A119 | Not identified | Poaceascoma sp. | Poaceascoma af. halophilum | Poaceascoma sp. | Lentitheciaceae | |
| A122 | Not identified | Poaceascoma sp. | Poaceascoma af. halophilum | Poaceascoma sp. | ||
| A120 | Not identified | Poaceascoma sp. | Not identified | Poaceascoma sp. | ||
| A121 | Not identified | Poaceascoma sp. | Not identified | Poaceascoma sp. | ||
| A123 | Not identified | Poaceascoma sp. | Not identified | Poaceascoma sp. | ||
| Class Leotiomycetes | ||||||
| A118 | Phialocephala sp. | Phialocephala sp. | Phialocephala af. bamuru | Phialocephala sp. | Vibrisseaceae | Helotiales |
| Class Sordariomycetes | ||||||
| A114 | Nigrospora sp. | Not identified | Not identified | Nigrospora sp. | Apiosporaceae | Xylariales |
| A116 | Nigrospora sp. | Nigrospora zimmermanii | Nigrospora zimmermanii | Nigrospora zimmermanii | Apiosporaceae | |
| A117 | Nigrospora sp. | Not identified | Not identified | Nigrospora sp. | Apiosporaceae | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fors, R.O.; Patreze, C.M.; Louro Berbara, R.L.; Carbone Carneiro, M.A.; Saggin-Júnior, O.J. Dark Septate Endophytic Fungi Associated with Sugarcane Plants Cultivated in São Paulo, Brazil. Diversity 2020, 12, 351. https://doi.org/10.3390/d12090351
Fors RO, Patreze CM, Louro Berbara RL, Carbone Carneiro MA, Saggin-Júnior OJ. Dark Septate Endophytic Fungi Associated with Sugarcane Plants Cultivated in São Paulo, Brazil. Diversity. 2020; 12(9):351. https://doi.org/10.3390/d12090351
Chicago/Turabian StyleFors, Rosalba Ortega, Camila Maistro Patreze, Ricardo Luis Louro Berbara, Marco Aurélio Carbone Carneiro, and Orivaldo José Saggin-Júnior. 2020. "Dark Septate Endophytic Fungi Associated with Sugarcane Plants Cultivated in São Paulo, Brazil" Diversity 12, no. 9: 351. https://doi.org/10.3390/d12090351