
Dry Rot Caused by the Complex Colletotrichum falcatum and Thielaviopsis paradoxa Emerges as a Key Stalk Disorder in Newly Expanded Sugarcane Plantations from Northwestern São Paulo, Brazil

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Survey of Sugarcane Stalk-Related Disorders
2.2. Mapping and Estimates Related to the Field Prevalence and Distribution of Three Stalk-Associated Disorders in Sugarcane Plantations
2.2.1. Mapping the Frequency Field Distribution of Sugarcane Stalk-Associated Disorders in Northwestern São Paulo
2.2.2. Estimating the Prevalence and Distribution of Sugarcane Stalk-Associated Disorders
- (1)
- TL = (Estimated yield–Actual yield)/total area sampled, where TL is total yield loss in the area in t·ha−1;
- (2)
- RL = [(Estimated yield–Actual yield)/Estimated yield], where RL is the relative yield loss in each area as a percentage;
- (3)
- rrTL = TL × [(I.rr)/(I.rr + I.sib + I.dr)], where rrTL is the yield loss associated with the incidence of the red rot disease (t·ha−1), I.rr = incidence of the red rot disease (%), I.sib = incidence of the spittlebug-induced budding (%), and I.dr = incidence of the stalk internal dry rot (%);
- (4)
- rrRL = RL × [(I.rr)/(I.rr + I.sib + I.dr)], where rrRL is the relative yield loss associated with the red rot disease (%);
- (5)
- sbiTL and (6) drTL (the yield losses associated with spittlebug-induced budding or stalk internal dry rot) were calculated in a similar manner to (3), while (7) sbiRL and (8) drRL (the corresponding relative yield losses) were calculated in a similar manner to (4).
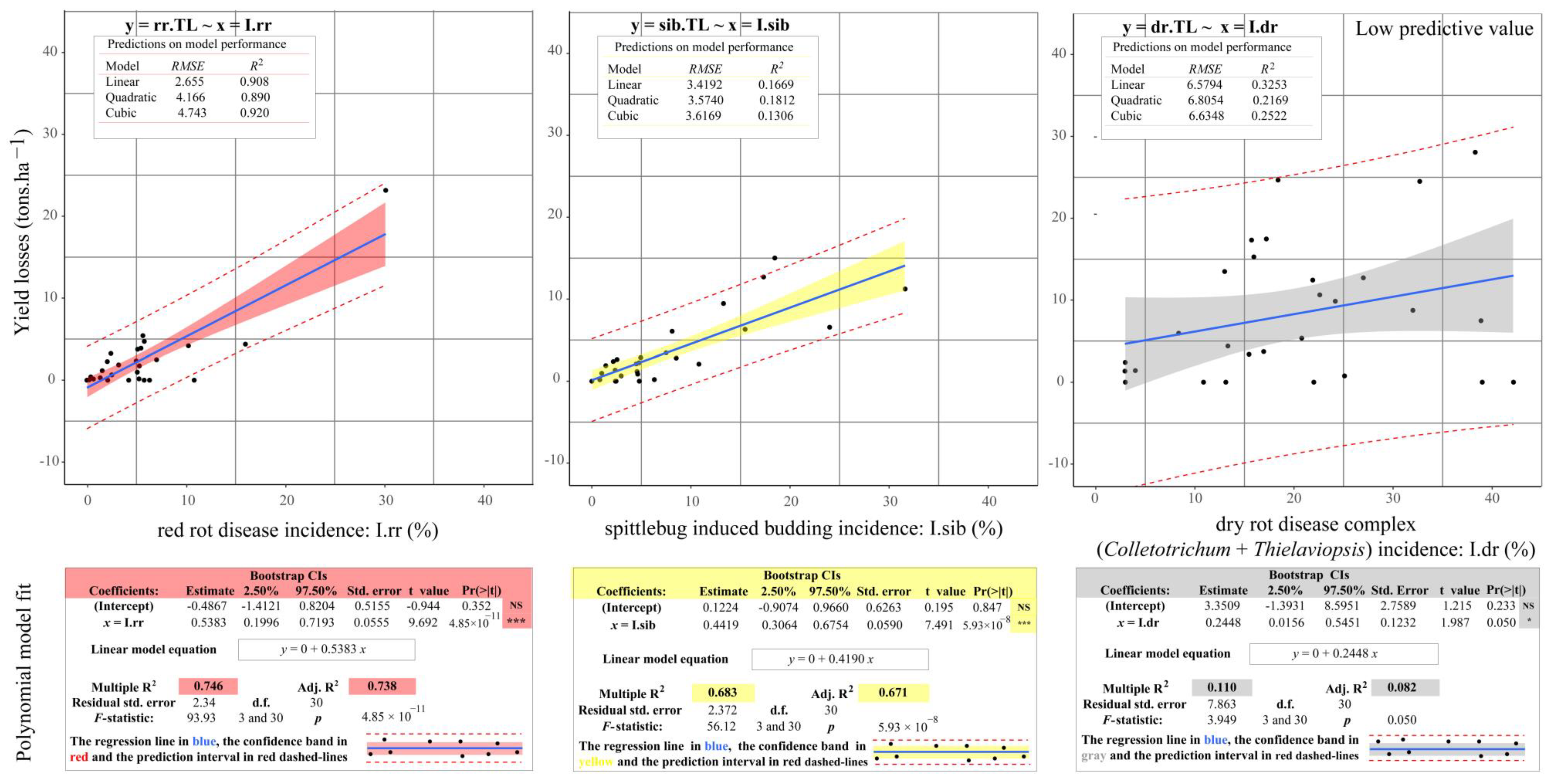
2.3. Predicting the Yield Losses in Northwestern São Paulo Sugarcane Fields on the Basis of Each Independent Stalk-Related Disorder
| BootstrapRandomX <- function(dat=mydata, mod.formula=formula (TL~rrTL + sbiTL + drTL)) {dat.boot <- dat[sample(x = nrow(dat), size = nrow(dat), replace=T),] boot.lm <- lm(mod.formula, data=dat.boot) coef(boot.lm)} N = 5000 vector.boot <- t(replicate(N, BootstrapRandomX())) apply(vector.boot, MARGIN = 2, sd) t(apply(vector.boot, MARGIN = 2, quantile, probs=c(0.025, 0.975))) |
| # Building the model–Linear model: model <- lm(y~x, data = train.data) # For a quadric or cubic polynomial, the model would be: model <- lm(y~poly(x, 2, raw = TRUE), data = train.data) # for quadratic model <- lm(y~poly(x, 3, raw = TRUE), data = train.data) # for cubic # Making predictions: predictions <- model %>% predict(test.data) # Model performance data.frame(RMSE = RMSE(predictions, test.data$y), + R2 = R2(predictions, test.data$y)) # Visualizing the data: p1<-ggplot(train.data, aes(x, y)) + geom_point() + stat_smooth(method = lm, formula = y~x) p1 # For a quadric or cubic polynomial, the code would be: stat_smooth(method = lm, formula = y~poly(x, 2, raw = TRUE)) # for quadratic stat_smooth(method = lm, formula = y~poly(x, 3, raw = TRUE)) # for cubic # Adding predictions: pred.int <- predict(model, interval = “prediction”) datalwrupr <- cbind(train.data, pred.int) # Regression line plus confidence intervals: p2 <- ggplot(datalwrupr, aes(x, y)) + geom_point() + coord_cartesian(xlim = c(0, 45), ylim = c(-10,45))+ stat_smooth(method = lm, formula = y~x) + # For a quadric or cubic polynomial, the code would be: stat_smooth(method = lm, formula = y~poly(x, 2, raw = TRUE)) + # for quadratic stat_smooth(method = lm, formula = y~poly(x, 3, raw = TRUE)) + # for cubic # Adding prediction intervals to the regression line: geom_line(aes(y = lwr), color = “red”, linetype = “dashed”)+ geom_line(aes(y = upr), color = “red”, linetype = “dashed”)+ theme (panel.background = element_rect(fill = “white”, color = “black”), panel.grid.minor = element_line(color = “gray50”), panel.spacing = unit(5, “lines”)) p2 |
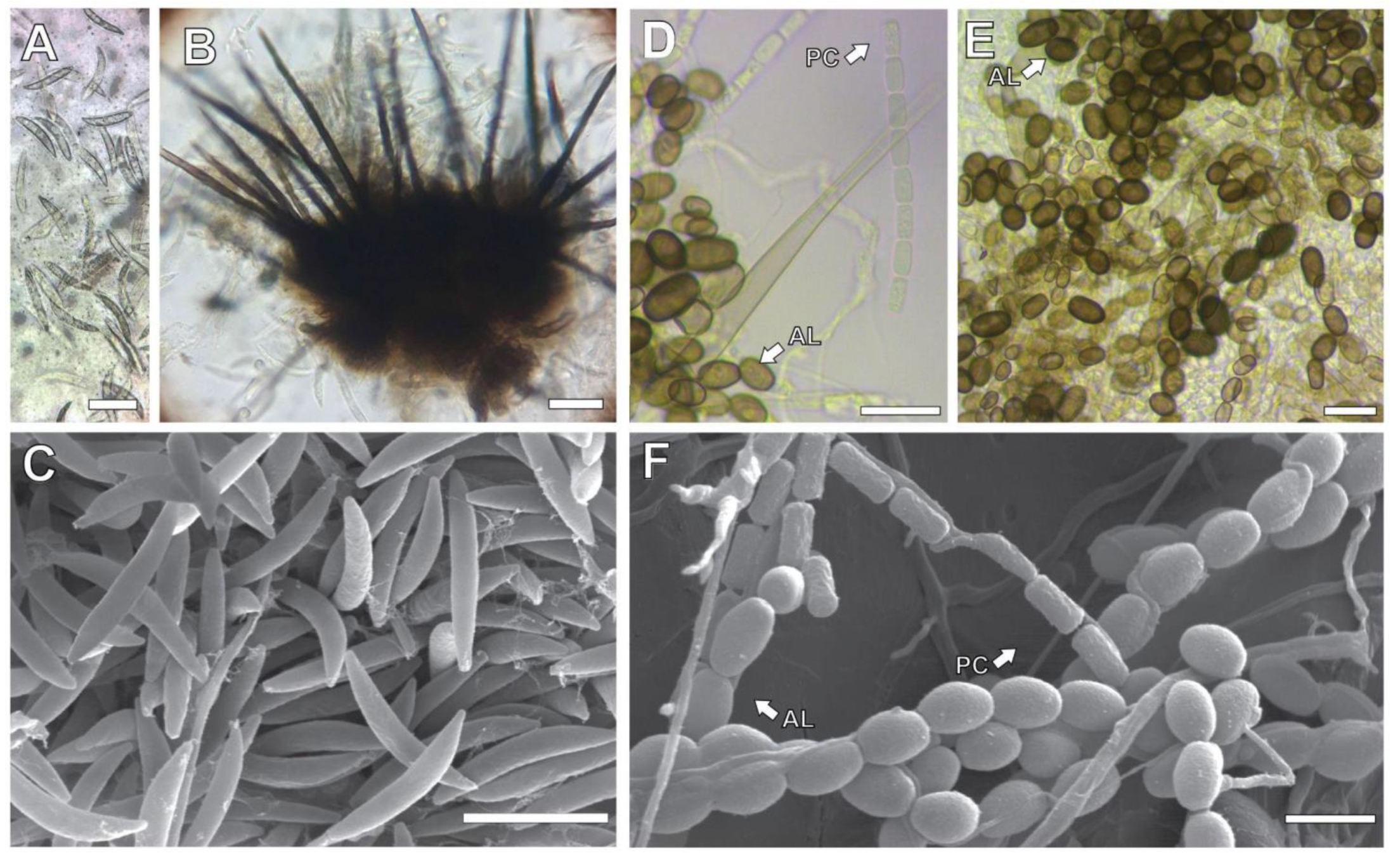
2.4. Detection of Pathogens Associated with Sugarcane Stalk Internal Dry Rot
3. Results
3.1. Mapping the Frequency Field Distribution of Sugarcane Stalk-Associated Disorders in Northwestern São Paulo
3.2. Estimating the Prevalence and Distribution of Sugarcane Stalk-Associated Disorders
3.3. Predicting Yield Losses in Northwestern São Paulo Sugarcane Fields on the Basis of Each Independent Stalk-Related Disorder
3.4. Detection of Pathogens Associated with Sugarcane Stalk Internal Dry Rot
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Companhia Nacional de Abastecimento (Conab). Acompanhamento Da Safra Brasileira de Cana-de-Açúcar, Safra 2022/23, 4o. Levantamento; Conab: Brasília, DF, Brazil, 2023; Volume 10. [Google Scholar]
- Cortez, L.A.B. Sugarcane Bioethanol: R & D for Productivity and Sustainability; Edgard Blucher: São Paulo, Brazil, 2010; ISBN 978-85-212-0822-8. [Google Scholar]
- Baeninger, R.; Peres, R.G. Regiões Canavieiras; Por dentro do estado de São Paulo; Núcleo de Estudos de População (Nepo)—Unicamp: Campinas, SP, Brazil, 2013; ISBN 978-85-88258-39-6. [Google Scholar]
- Hespanhol, A.N. Dinâmica Agroindustrial, Intervenção Estatal e a Questão do Desenvolvimento da Região de Andradina. Ph.D. Dissertation, São Paulo State University (UNESP), Rio Claro, SP, Brazil, 1996. [Google Scholar]
- Leite, R.C.D.C.; Leal, M.R.L.V. O Biocombustível No Brasil. Novos Estud. CEBRAP 2007, 15–21. [Google Scholar] [CrossRef]
- Landau, E.C. Dinâmica da Produção Agropecuária e da Paisagem Natural no Brasil nas Últimas Décadas; Embrapa: Brasília, DF, Brazil, 2020; Volume 2, ISBN 9786587380032. [Google Scholar]
- Demétrio, N.B. Novas áreas de expansão da cana-de-açúcar no Oeste Paulista: Do agrário ao território da produção internacional de commodities. In Regiões Canavieiras; Por Dentro do Estado de São Paulo; Núcleo de Estudos de População (Nepo)—Unicamp: Campinas, SP, Brazil, 2013; pp. 119–140. ISBN 978-85-88258-39-6. [Google Scholar]
- Lopes, F.S.; Ribeiro, H. Mapping of Hospitalizations Due to Respiratory Problems and Possible Associations to Human Exposure to Burnt Sugar-Cane Straw Products in the State of São Paulo. Rev. Bras. Epidemiol. 2006, 9, 215–225. [Google Scholar] [CrossRef]
- Ribeiro, H. Sugar Cane Burning in Brazil: Respiratory Health Effects. Rev. Saúde Pública 2008, 42, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Cassila, J.C.; Marcel, F. Conflito de Competência Legislativa Em Matéria Ambiental e a Queima Da Palha Da Cana-de-Açúcar: Uma Análise Da Jurisprudência Do Tribunal de Justiça Do Estado de São Paulo. Rev. Eletrônica FACP 2014, 5, 51–73. [Google Scholar]
- Moura, L.; Landau, E. Evolução da produção da cana-de-açúcar. In Dinâmica da Produção Agropecuária e da Paisagem Natural no Brasil nas Últimas Décadas; Embrapa: Brasília, DF, Brazil, 2020; Volume 2, pp. 625–650. ISBN 978-65-87380-03-2. [Google Scholar]
- Legislative Assembly of the State of São Paulo Decree 47700, from March 11, 2003: Regulates Law No. 11241, from September 19, 2002, Which Provides for the Gradual Elimination of the Burning of Sugarcane Straw and Provides Related Measures. Diário Estado—Poder Exec, 180th ed.; 2023; Section 1, p. 2.
- INPE—Instituto Nacional de Pesquisa Espaciais—Brazil. Canasat. 2012. Available online: http://www.dsr.inpe.br/laf/canasat/colheita.html (accessed on 2 May 2023).
- Dorneles Junior, J.; Alves, R.P.; Santos, R.M.; Ramos, R.; Ramos, N.P.; Prado, S.S. Influência da quantidade de palhada em cana-de-açúcar na população de Mahanarva fimbriolata (Stâl, 1854) (Hemiptera: Cercopidae). In Workshop Agroenergia: Matérias Primas, 9., 2015, Ribeirão Preto, Anais; APTA IAC: Ribeirão Preto, SP, Brazil, 2015; pp. 7–88. [Google Scholar]
- Macedo, N.; Campos, M.B.S.; Araújo, J.R. Insetos Nas Raízes e Colo Da Planta, Perfilhamento e Produtividade Em Canaviais Colhidos Com e Sem Queima. Rev. Soc. Téc. Açucar. E Alcooleiros Bras. 1997, 15, 18–21. [Google Scholar]
- Nechet, K.L.; Ramos, N.P.; Halfeld-Vieira, B.A. Identificação de Doenças Fúngicas Foliares Emergentes Em Cana-de-Açúcar. In Comunicado Técnico; Embrapa Meio Ambiente: Jaguariúna, Brazil, 2016; p. 5. ISBN 1516-8638. [Google Scholar]
- Silva, Y.T. Sugarcane Spittlebug (Mahanarva fimbriolata): A Review of the Species Aspects, Its Damage to Sugarcane Cultivation and Perspectives of Biological Control. Bachelor’s Thesis, Universidade Federal de São Carlos (UFSCar), Araras, Brazil, 2022. [Google Scholar]
- Macedo, N.; Araújo, J.R. Efeitos Da Queima Do Canavial Sobre Insetos Predadores. An. Soc. Entomológica Bras. 2000, 29, 71–77. [Google Scholar] [CrossRef]
- Buriozo, G.S.; Caramelo, A.D.; Campos, A.P. Perfilhamento e Ocorrência de Cigarrinha Da Raiz Em Lavouras de Cana-de-Açúcar Em Sistemas de Colheita Com e Sem Palha. Sci. Technol. Innov. Agron. 2017, 1, 46–53. [Google Scholar]
- Marins, É.F.C.; Da Silva, M.J.D.S.; Da Silva, J.L.; Silva-Cabral, J.R.A.; Costa, J.F.D.O.; Feijó, F.M.; Assunção, I.P.; Lima, G.S.d.A. Colletotrichum Species Associated with Sugarcane Red Rot in Brazil. Fungal Biol. 2022, 126, 290–299. [Google Scholar] [CrossRef]
- Abbott, E.V. Red Rot of Sugarcane. In Technical Bulletin; USDA: Washington, DC, 1938; p. 96. [Google Scholar]
- Costa, M.M.; Silva, B.A.A.S.; Moreira, G.M.; Pfenning, L.H. Colletotrichum Falcatum and Fusarium Species Induce Symptoms of Red Rot in Sugarcane in Brazil. Plant Pathol. 2021, 70, 1807–1818. [Google Scholar] [CrossRef]
- Viswanathan, R. Plant Disease: Red Rot of Sugarcane; Anmol Publications: New Delhi, India, 2010; ISBN 978-81-261-4214-9. [Google Scholar]
- Borges, A.F.; De Alcântara Neto, F.; Da Silva Matos, K.; Júnior, J.E.A.B.; Júnior, N.S.M.; Moreira, S.I.; De Melo, M.P. Thielaviopsis Ethacetica the Etiological Agent of Sugarcane Pineapple Sett Rot Disease in Brazil. Trop. Plant Pathol. 2019, 44, 460–467. [Google Scholar] [CrossRef]
- Sansoli Chanquinie, D.M. Pathogenic Diversity of Thielaviopsis paradoxa Isolates from Different Sugarcane Producing Areas. Master’s Thesis, São Paulo State University (UNESP), Jaboticabal, SP, Brazil, 2015. [Google Scholar]
- Raid, R.N. Pineapple Disease of Sugarcane; University of Florida, Everglades Research and Education Center: Belle Glade, FL, USA, 2012. [Google Scholar]
- Aparecido, C.C.; Azania, C.A.M. Podridão Abacaxi Em Cana-de-Açúcar; Comunicado técnico 175; Instituto Biológico: São Paulo, Brazil, 2012. [Google Scholar]
- Center for Agriculture and Bioscience International (CABI). Ceratocystis Paradoxa (Black Rot of Pineapple); CABI: Wallingford, UK, 2022; p. 12157. [Google Scholar] [CrossRef]
- Tokeshi, H.; Rago, A. Doenças Da Cana-de-Açúcar. In Manual de Fitopatologia: Doenças das Plantas Cultivadas; Ceres: São Paulo, Brazil, 2005; Volume 2, pp. 185–196. [Google Scholar]
- Chapola, R.G.; Ogasawara, G.A.; Jans, B.; Massola Junior, N.S. Control of Pineapple Disease of Sugarcane through In-Furrow Fungicide Spray Application on Seedpieces. Ciênc. Rural 2014, 44, 197–202. [Google Scholar] [CrossRef]
- Matsuoka, S. Manejo de Doenças e Medidas de Controle. In Cana-de-Açúcar: Do Plantio à Colheita; UFV: Viçosa, Brazil, 2016; pp. 108–138. [Google Scholar]
- Uznan, J. Sugarcane Pineapple Disease: Cultivars Reaction, Chemical Control and Pathogenic Diversity. Master’s Thesis, Universidade Federal de São Carlos (UFSCar), Araras, Brazil, 2019. [Google Scholar]
- Garcia, D.B.; Ravaneli, G.C.; Madaleno, L.L.; Mutton, M.A.; Mutton, M.J.R. Damages of Spittlebug on Sugarcane Quality and Fermentation Process. Sci. Agric. 2010, 67, 555–561. [Google Scholar] [CrossRef]
- Parra, J.R.; Botelho, R.; Pinto, A.S. Biological control of pests as a key component for sustainable sugarcane production. In Sugarcane Bioethanol: R e D for Productivity and Sustainability; Editora Edgard Blucher: São Paulo, Brazil, 2010; pp. 441–450. ISBN 978-85-212-0822-8. [Google Scholar]
- Lopes, D.O.P. Performance of Sugarcane Genotypes in Relation to the Borer-Rot Complex Caused by Diatraea Saccharalis (Fabricius, 1794) (Lepidoptera: Crambidae) and Microorganisms. Master’s Thesis, São Paulo State University (UNESP), Jaboticabal, SP, Brazil, 2012. [Google Scholar]
- Medeiros, C.C.P. Genetic Diversity of Thielaviopsis Paradoxa Isolates from Sugarcane. Master’s Thesis, São Paulo State University (UNESP), Jaboticabal, SP, Brazil, 2015. [Google Scholar]
- Ferreira, M.D.C.; Werneck, C.F.; Furuhashi, S.; Leite, G.J. Tratamento de Toletes de Cana-de-Açúcar Para o Controle Da Podridão-Abacaxi Em Pulverização Conjugada Ao Plantio Mecanizado. Eng. Agríc. 2008, 28, 263–273. [Google Scholar] [CrossRef]
- Almeida, J.E.M.; Batista Filho, A.; Santos, A.S. Avaliação Do Controle Biológico de Mahanarva Fimbriolata (Hom., Cercopidae) Com o Fungo Metarhizium Anisopliae Em Variedades de Cana-de-Açúcar e Diferentes Épocas de Corte. Arq. Inst. Biológico 2003, 70, 101–103. [Google Scholar]
- Almeida, J.E.M.; Batista Filho, A. Controle Biológico de Cigarrinha Da Raiz Da Cana-de-Açúcar; Tecnologia Sustentável; Instituto Biológico: São Paulo, Brazil, 2006. [Google Scholar]
- Braga, N.C.D.C.; Severiano, E.D.C.; Santos, L.D.S.; Rúbio Neto, A.; Rodrigues, T.M.; Lima, J.D.P. Production of Sugarcane Seedlings Pre-Sprouted in Commercial and Alternative Substrates with by-Products of the Sugarcane Industry. Semina Ciênc. Agrár. 2019, 40, 33. [Google Scholar] [CrossRef]
- Oliveira, M.H.R. Production of Pre-Sprouted Sugarcane Seedlings under Localized Irrigation. Master’s Thesis, Instituto Federal de Educação, Ciência e Tecnologia Goiano (IFGO), Ceres, Goiás, Brazil, 2019. [Google Scholar]
- Santos, L.S.; Braga, N.C.C.; Rodrigues, T.M.; Neto, A.R.; Brito, M.F.; Da Costa Severiano, E. Pre-Sprouted Seedlings of Sugarcane Using Sugarcane Industry by-Products as Substrate. Sugar Tech 2020, 22, 675–685. [Google Scholar] [CrossRef]
- Brandi, F.; Heck, D.W.; Ferreira, T.C.; Bettiol, W. Commercial Formulations of Bacillus Spp. for Sugarcane Pineapple Disease Control and Growth Promotion. Pesqui. Agropecuária Bras. 2018, 53, 1311–1319. [Google Scholar] [CrossRef]
- Cursi, D.E.; Hoffmann, H.P.; Barbosa, G.V.S.; Bressiani, J.A.; Gazaffi, R.; Chapola, R.G.; Fernandes Junior, A.R.; Balsalobre, T.W.A.; Diniz, C.A.; Santos, J.M.; et al. History and Current Status of Sugarcane Breeding, Germplasm Development and Molecular Genetics in Brazil. Sugar Tech 2022, 24, 112–133. [Google Scholar] [CrossRef]
- Borges Filho, R.D.C.; Sturza, V.S.; Bernardi, D.; Da Cunha, U.S.; Pinto, A.S.; Dos Anjos E Silva, S.D.; Nava, D.E. Population Dynamics of Pests and Natural Enemies on Sugar Cane Grown in a Subtropical Region of Brazil. Fla. Entomol. 2019, 102, 526. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Bruce, P.C.; Bruce, A.; Gedeck, P. Practical Statistics for Data Scientists: 50+ Essential Concepts Using R and Python, 2nd ed.; O’Reilly Media, Inc.: Sebastopol, CA, USA, 2020; ISBN 978-1-4920-7289-8. [Google Scholar]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill Book Company, Inc.: London, UK, 1940. [Google Scholar]
- Bozzola, J.J.; Russell, L.D. Electron Microscopy, 2nd ed.; Jones and Bartlett Publishers: Boston, MA, USA, 1999. [Google Scholar]
- Hossain, M.I.; Ahmad, K.; Vadamalai, G.; Siddiqui, Y.; Saad, N.; Ahmed, O.H.; Hata, E.M.; Adzmi, F.; Rashed, O.; Rahman, M.Z.; et al. Phylogenetic Analysis and Genetic Diversity of Colletotrichum Falcatum Isolates Causing Sugarcane Red Rot Disease in Bangladesh. Biology 2021, 10, 862. [Google Scholar] [CrossRef]
- Bernardo, R.; Lourenzani, W.L.; Satolo, E.G.; Caldas, M.M. Analysis of the Agricultural Productivity of the Sugarcane Crop in Regions of New Agricultural Expansions of Sugarcane. Gest. Produção 2019, 26, e3554. [Google Scholar] [CrossRef]
- Agnihotri, V.P. Diseases of Sugarcane and Sugarbeet; Oxford & IBH Publ: New Delhi, India, 1990; ISBN 978-81-204-0511-0. [Google Scholar]
- Seeja, G.; Sreekumar, K. Character Association between Yield and Its Related Traits Governing Redrot Disease Incidence in Inter Varietal Crosses of Sugarcane. Agric. Sci. Dig. 2009, 29, 24–29. [Google Scholar]
- Meyer, J.; Rein, P.; Turner, P.; Mathias, K. Good Management Practices Manual for the Cane Sugar Industry; PGBI Sugar & Bioenergy (Pty) Ltd., International Finance Corporation (IFC): Johannesburg, South Africa, 2011. [Google Scholar]
- Almeida Neto, L.A.; Guiselini, C.; Menezes, D.; Cordeiro Júnior, J.J.F.; Pandorfi, H. Growth of Pre-Sprouted Sugarcane Seedlings Submitted to Supplementary Lighting. Rev. Bras. Eng. Agríc. E Ambient 2020, 24, 194–199. [Google Scholar] [CrossRef]
- Franco, H.C.J.; Pimenta, M.T.B.; Carvalho, J.L.N.; Magalhães, P.S.G.; Rossell, C.E.V.; Braunbeck, O.A.; Vitti, A.C.; Kölln, O.T.; Rossi Neto, J. Assessment of Sugarcane Trash for Agronomic and Energy Purposes in Brazil. Sci. Agric. 2013, 70, 305–312. [Google Scholar] [CrossRef]
- Gonzaga, L.C.; Zotelli, L.D.C.; De Castro, S.G.Q.; De Oliveira, B.G.; Bordonal, R.D.O.; Cantarella, H.; Carvalho, J.L.N. Implications of Sugarcane Straw Removal for Soil Greenhouse Gas Emissions in São Paulo State, Brazil. BioEnergy Res. 2019, 12, 843–857. [Google Scholar] [CrossRef]
- Vitti, A.C.; Franco, H.C.J.; Trivelin, P.C.O.; Ferreira, D.A.; Otto, R.; Fortes, C.; Faroni, C.E. Nitrogênio Proveniente Da Adubação Nitrogenada e de Resíduos Culturais Na Nutrição Da Cana-Planta. Pesqui. Agropecuária Bras. 2011, 46, 287–293. [Google Scholar] [CrossRef]
- Fortes, C.; Trivelin, P.C.O.; Vitti, A.C. Long-Term Decomposition of Sugarcane Harvest Residues in Sao Paulo State, Brazil. Biomass Bioenergy 2012, 42, 189–198. [Google Scholar] [CrossRef]
- Dominick, G.; Avanci, C.E.; Silva Júnior, D.R.; Bueno, E.F.; Pereira Filho, F.; Macedo, J.O.R.; Cintra, G.S. Comportamento de variedades e patogenicidade de fungos associados à podridões em cana-de-açúcar. In A Face Transdisciplinar das Ciências Agrárias, 2; Atena: Ponta Grossa, Paraná, Brazil, 2021; pp. 42–55. [Google Scholar]
- McDonald, B.A.; Linde, C. Pathogen Population Genetics, Evolutionary Potential, and Durable Resistance. Annu. Rev. Phytopathol. 2002, 40, 349–379. [Google Scholar] [CrossRef]
- McDonald, B.A.; Stukenbrock, E.H. Rapid Emergence of Pathogens in Agro-Ecosystems: Global Threats to Agricultural Sustainability and Food Security. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2016, 371, 20160026. [Google Scholar] [CrossRef]
- Singh, V.; Srivastava, S.N.; Lal, R.J.; Awasthi, S.K.; Joshi, B.B. Biological Control of Red Rot Disease of Sugarcane through Trichoderma Harzianum and Trichoderma Viride. Indian Phytopathol. 2008, 61, 486–691. [Google Scholar]
- MAPA—Ministério da Agricultura Pecuária e Abastecimento—Brazil. Agrofit—Sistemas de Agrotóxicos Fitossanitários, Coordenação Geral de Agrotóxicos e Afins. 2023. Available online: http://agrofit.agricultura.gov.br/agrofit_cons/principal_agrofit_cons (accessed on 2 May 2023).
- Palmieri, D.; Ianiri, G.; Del Grosso, C.; Barone, G.; De Curtis, F.; Castoria, R.; Lima, G. Advances and Perspectives in the Use of Biocontrol Agents against Fungal Plant Diseases. Horticulturae 2022, 8, 577. [Google Scholar] [CrossRef]
- Iwanicki, N.S.; Pereira, A.A.; Botelho, A.B.R.Z.; Rezende, J.M.; Moral, R.D.A.; Zucchi, M.I.; Delalibera Júnior, I. Monitoring of the Field Application of Metarhizium anisopliae in Brazil Revealed High Molecular Diversity of Metarhizium Spp. in Insects, Soil and Sugarcane Roots. Sci. Rep. 2019, 9, 4443. [Google Scholar] [CrossRef] [PubMed]
- Segato, S.V.; Pinto, A.S.; Jendiroba, E.; Nóbrega, J.C.M. Atualização em Produção de Cana-de-Açúcar; CP 2: Piracicaba, SP, Brazil, 2006. [Google Scholar]
- Vuyyuru, M.; Sandhu, H.; McCray, J.; Raid, R. Effects of Soil-Applied Fungicides on Sugarcane Root and Shoot Growth, Rhizosphere Microbial Communities, and Nutrient Uptake. Agronomy 2018, 8, 223. [Google Scholar] [CrossRef]
- Dinardo-Miranda, L.L.; Garcia, V.; Parazzi, V.J. Efeito de Inseticidas No Controle de Mahanarva Fimbriolata (Stål) (Hemiptera: Cercopidae) e de Nematóides Fitoparasitos Na Qualidade Tecnológica e Na Produtividade Da Cana-de-Açúcar. Neotrop. Entomol. 2002, 31, 609–614. [Google Scholar] [CrossRef]
- Espitia Buitrago, P.A.; Manzano, M.R.; Hernández, L.M. Spittlebugs (Hemiptera: Cercopidae): Integrated Pest Management on Gramineous Crops in the Neotropical Ecozone. Front. Sustain. Food Syst. 2022, 6, 891417. [Google Scholar] [CrossRef]
- Hollomon, D.W. Fungicide Resistance: 40 Years on and Still a Major Problem. In Fungicide Resistance in Plant Pathogens; Ishii, H., Hollomon, D.W., Eds.; Springer: Tokyo, Japan, 2015; pp. 3–11. ISBN 978-4-431-55641-1. [Google Scholar]
- Ceresini, P.C.; Silva, T.C.; Vicentini, S.N.C.; Leite, R.P., Jr.; Moreira, S.I.; Castro-Ríos, K.; Garcés-Fiallos, F.R.; Krug, L.D.; Moura, S.S.; Silva, A.G.; et al. Strategies for Managing Fungicide Resistance in the Brazilian Tropical Agroecosystem: Safeguarding Food Safety, Health, and the Environmental Quality. Trop. Plant Pathol. 2023. accepted. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| County and Sugarcane Variety | Cropped Area Surveyed (ha) | Stalk-Related Disorder Incidence (%) | Total Losses in the Areas (t) | Yield Losses (TL) (t·ha−1) | Relative Yield Losses (RL) (%) | Yield Losses Associated with the Stalk Disorders (t·ha−1) | Relative Yield Losses Associated with the Stalk Disorders (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Red Rot (I.rr) | Spittlebug (I.sib) | Dry Rot (I.dr) | Red Rot (rrTL) | Spittlebug (sbiTL) | Dry Rot (drTL) | Red Rot (rrRL) | Spittlebug (sbiRL) | Dry Rot (drRL) | |||||
| Castilho, SP81-3250 | 151.32 | 12.68 | 11.00 | 25.00 | 1938.62 | 12.81 | 19.71 | 3.34 | 2.89 | 6.58 | 5.14 | 4.45 | 10.12 |
| 258.26 | 30.16 | 12.34 | 0.00 | 8427.94 | 32.63 | 42.04 | 23.16 | 9.47 | 0.00 | 29.83 | 12.21 | 0.00 | |
| 333.79 | 3.20 | 4.93 | 30.55 | 7499.61 | 22.47 | 34.57 | 1.86 | 2.87 | 17.74 | 2.86 | 4.41 | 27.30 | |
| Ilha Solteira, CTC4 | 78.42 | 0.00 | 12.00 | 39.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 95.27 | 16.00 | 24.00 | 32.00 | 1875.56 | 19.69 | 23.27 | 4.37 | 6.56 | 8.75 | 5.17 | 7.76 | 10.34 | |
| Ilha Solteira, SP81-3250 | 216.10 | 2.42 | 1.41 | 18.39 | 6438.41 | 29.79 | 49.66 | 3.25 | 1.89 | 24.66 | 5.41 | 3.15 | 41.10 |
| 420.46 | 30.91 | 7.50 | 2.94 | 7988.70 | 19.00 | 25.33 | 14.20 | 3.44 | 1.35 | 18.94 | 4.59 | 1.80 | |
| Itapura, SP81-3250 | 47.95 | 0.39 | 2.54 | 13.00 | 792.72 | 16.53 | 15.03 | 0.41 | 2.64 | 13.49 | 0.37 | 2.40 | 12.26 |
| 104.93 | 2.51 | 4.57 | 20.78 | 753.62 | 7.18 | 11.97 | 0.65 | 1.18 | 5.36 | 1.08 | 1.96 | 8.93 | |
| 157.28 | 0.00 | 4.48 | 27.00 | 2332.28 | 14.83 | 19.77 | 0.00 | 2.11 | 12.72 | 0.00 | 2.81 | 16.96 | |
| 268.32 | 2.06 | 7.02 | 16.00 | 7394.32 | 27.56 | 22.96 | 2.26 | 7.71 | 17.58 | 1.89 | 6.42 | 14.65 | |
| 325.07 | 4.51 | 2.17 | 15.72 | 8025.85 | 24.69 | 32.92 | 4.97 | 2.39 | 17.33 | 6.63 | 3.19 | 23.10 | |
| 405.36 | 5.46 | 13.29 | 8.36 | 7821.90 | 19.30 | 25.73 | 3.88 | 9.46 | 5.95 | 5.18 | 12.61 | 7.94 | |
| Pereira Barreto, SP81-3250 | 15.20 | 6.32 | 4.79 | 42.15 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 21.00 | 4.22 | 0.00 | 17.62 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 66.00 | 7.03 | 31.62 | 3.99 | 999.78 | 15.15 | 21.64 | 2.50 | 11.23 | 1.42 | 3.57 | 16.05 | 2.03 | |
| 127.79 | 10.25 | 15.47 | 24.17 | 2599.85 | 20.34 | 29.06 | 4.18 | 6.31 | 9.86 | 5.97 | 9.01 | 14.08 | |
| 131.56 | 5.79 | 18.47 | 2.97 | 2912.99 | 22.14 | 29.52 | 4.71 | 15.02 | 2.42 | 6.28 | 20.02 | 3.22 | |
| 117.51 | 10.83 | 2.45 | 22.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| S.A.Aracanguá, SP81-3250 | 25.07 | 0.66 | 2.96 | 15.46 | 104.87 | 4.18 | 7.61 | 0.14 | 0.65 | 3.39 | 0.26 | 1.18 | 6.16 |
| 33.90 | 1.34 | 0.81 | 16.94 | 141.96 | 4.19 | 7.61 | 0.29 | 0.18 | 3.72 | 0.54 | 0.32 | 6.76 | |
| 75.03 | 0.00 | 2.36 | 21.89 | 1033.95 | 13.78 | 25.06 | 0.00 | 1.34 | 12.44 | 0.00 | 2.44 | 22.62 | |
| 123.88 | 0.33 | 1.95 | 17.21 | 2449.37 | 19.77 | 32.95 | 0.33 | 1.98 | 17.47 | 0.55 | 3.29 | 29.11 | |
| 139.26 | 1.55 | 8.11 | 32.69 | 4419.05 | 31.73 | 45.33 | 1.16 | 6.07 | 24.50 | 1.66 | 8.68 | 35.00 | |
| 169.20 | 4.98 | 4.76 | 22.60 | 2574.46 | 15.22 | 23.41 | 2.34 | 2.24 | 10.64 | 3.60 | 3.44 | 16.36 | |
| 195.33 | 5.09 | 10.81 | 38.87 | 2058.17 | 10.54 | 14.05 | 0.98 | 2.08 | 7.48 | 1.30 | 2.77 | 9.97 | |
| 221.59 | 5.12 | 17.33 | 38.29 | 9862.19 | 44.51 | 59.34 | 3.75 | 12.70 | 28.05 | 5.00 | 16.93 | 37.41 | |
| Sud Mennucci, SP81-3250 | 12.07 | 2.10 | 0.00 | 13.12 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 32.21 | 0.23 | 4.63 | 17.36 | 133.34 | 4.14 | 6.37 | 0.04 | 0.86 | 3.23 | 0.07 | 1.33 | 4.98 | |
| 44.43 | 5.65 | 1.00 | 15.95 | 961.54 | 21.64 | 36.07 | 5.41 | 0.96 | 15.27 | 9.02 | 1.60 | 25.46 | |
| 46.67 | 5.30 | 8.52 | 13.33 | 417.58 | 8.95 | 14.91 | 1.75 | 2.81 | 4.39 | 2.91 | 4.68 | 7.32 | |
| 47.77 | 5.78 | 0.00 | 10.87 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 66.45 | 0.00 | 2.38 | 2.98 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
| 278.39 | 5.26 | 6.31 | 25.10 | 311.85 | 1.12 | 10.05 | 0.16 | 0.19 | 0.77 | 1.44 | 1.73 | 6.88 | |
| Mean | 142.73 | 5.83 | 7.41 | 19.54 | 2713.84 | 14.23 | 20.17 | 2.65 | 3.45 | 8.13 | 3.67 | 4.69 | 11.82 |
| 95% CI | 37.68 | 2.43 | 2.42 | 3.68 | 1044.87 | 3.83 | 5.22 | 1.51 | 1.34 | 2.72 | 1.98 | 1.76 | 3.93 |
| Total | 4852.84 | - | - | - | 92,270.48 | 483.88 | - | 90.09 | 117.23 | 276.56 | - | - | - |
| County, Sample Size (= N Fields) | Mean Cropped Area Surveyed (ha) | Stalk-Related Disorder Incidence (%) | Total Losses in the Areas (t) | Yield Losses (TL) (t·ha−1) | Relative Yield Losses (RL) (%) | Yield Losses Associated with the Disorders (t·ha−1) | Relative Yield Losses Associated with the Disorders (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Red Rot (I.rr) * | Spittlebug (I.sib) | Dry Rot (I.dr) | Red Rot–rrTL * | Spittlebug (sbiTL) | Dry Rot (drTL) | Red Rot (rrRL) * | Spittlebug (sbiRL) | Dry Rot (drRL) | ||||||||
| Castilho, N = 3 | 247.79 | 15.35 | a | 9.42 | 18.52 | 5955.39 | 22.64 | 32.11 | 9.45 | a | 5.08 | 8.11 | 12.61 | a | 7.02 | 12.47 |
| Ilha Solteira, N = 4 | 202.56 | 12.33 | a | 11.23 | 23.08 | 4075.67 | 17.12 | 24.57 | 5.46 | a | 2.97 | 8.69 | 7.38 | a | 3.87 | 13.31 |
| Itapura, N = 6 | 218.15 | 2.49 | b | 5.68 | 16.81 | 4520.12 | 18.35 | 21.40 | 2.03 | b | 4.25 | 12.07 | 2.52 | b | 4.90 | 13.97 |
| Pereira Barreto, N = 6 | 79.84 | 7.41 | b | 12.13 | 18.82 | 1085.44 | 9.61 | 13.37 | 1.90 | b | 5.43 | 2.28 | 2.64 | b | 7.51 | 3.22 |
| S.A.Aracanguá, N = 8 | 122.91 | 2.38 | b | 6.13 | 25.49 | 2830.50 | 17.99 | 26.92 | 1.12 | b | 3.41 | 13.46 | 1.61 | b | 4.88 | 20.42 |
| Sud Mennucci, N = 7 | 75.43 | 3.47 | b | 3.26 | 14.10 | 260.62 | 5.12 | 9.63 | 1.05 | b | 0.69 | 3.38 | 1.92 | b | 1.33 | 6.38 |
| Mean | 142.73 | 5.83 | 7.98 | 19.54 | 2713.84 | 14.23 | 20.17 | 2.65 | 3.45 | 8.13 | 3.67 | 4.69 | 11.82 | |||
| 95% CI | 37.68 | 2.43 | 2.55 | 3.68 | 1044.87 | 3.83 | 5.22 | 1.51 | 1.34 | 2.72 | 1.98 | 1.76 | 3.93 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, T.C.; Moreira, S.I.; Felizardo, L.M.; Martins, R.M.; Suzuki, A.N.; Faria, G.A.; Montanari, R.; Ceresini, P.C. Dry Rot Caused by the Complex Colletotrichum falcatum and Thielaviopsis paradoxa Emerges as a Key Stalk Disorder in Newly Expanded Sugarcane Plantations from Northwestern São Paulo, Brazil. Agronomy 2023, 13, 2729. https://doi.org/10.3390/agronomy13112729
Silva TC, Moreira SI, Felizardo LM, Martins RM, Suzuki AN, Faria GA, Montanari R, Ceresini PC. Dry Rot Caused by the Complex Colletotrichum falcatum and Thielaviopsis paradoxa Emerges as a Key Stalk Disorder in Newly Expanded Sugarcane Plantations from Northwestern São Paulo, Brazil. Agronomy. 2023; 13(11):2729. https://doi.org/10.3390/agronomy13112729
Chicago/Turabian StyleSilva, Tatiane Carla, Silvino Intra Moreira, Lucas Menezes Felizardo, Renan Milani Martins, Aline Namie Suzuki, Gláucia Amorim Faria, Rafael Montanari, and Paulo Cezar Ceresini. 2023. "Dry Rot Caused by the Complex Colletotrichum falcatum and Thielaviopsis paradoxa Emerges as a Key Stalk Disorder in Newly Expanded Sugarcane Plantations from Northwestern São Paulo, Brazil" Agronomy 13, no. 11: 2729. https://doi.org/10.3390/agronomy13112729