
The Combination of Synoeca-MP Antimicrobial Peptide with IDR-1018 Stimulates Proliferation, Migration, and the Expression of Pro-Regenerative Genes in Both Human Skin Cell Cultures and 3D Skin Equivalents

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Maintenance
2.2. Peptide Acquisition and Characterization
2.3. Peptide Treatment
2.4. Cell Viability and Proliferation
2.5. Cell Migration Assay
2.6. Gene Expression Analysis
2.7. Human 3D Skin Equivalent Wound Models
2.8. Antimicrobial Assays for Inhibitory and Bactericidal Concentrations against S. aureus
2.9. Drug Combination-Adapted Checker-Board Assay
2.10. Statistical Analysis
3. Results
3.1. IDR-1018 Did Not Compromise the Antibacterial Effect of Synoeca-MP
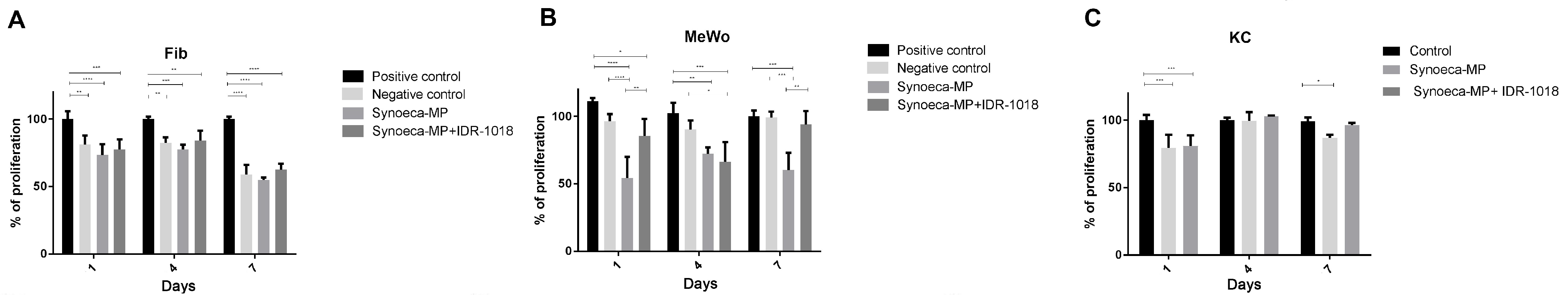
3.2. Cytotoxic Potential of Synoeca-MP on Skin Cells
3.3. IDR-1018 Peptide Reduces Synoeca-MP Cytotoxicity
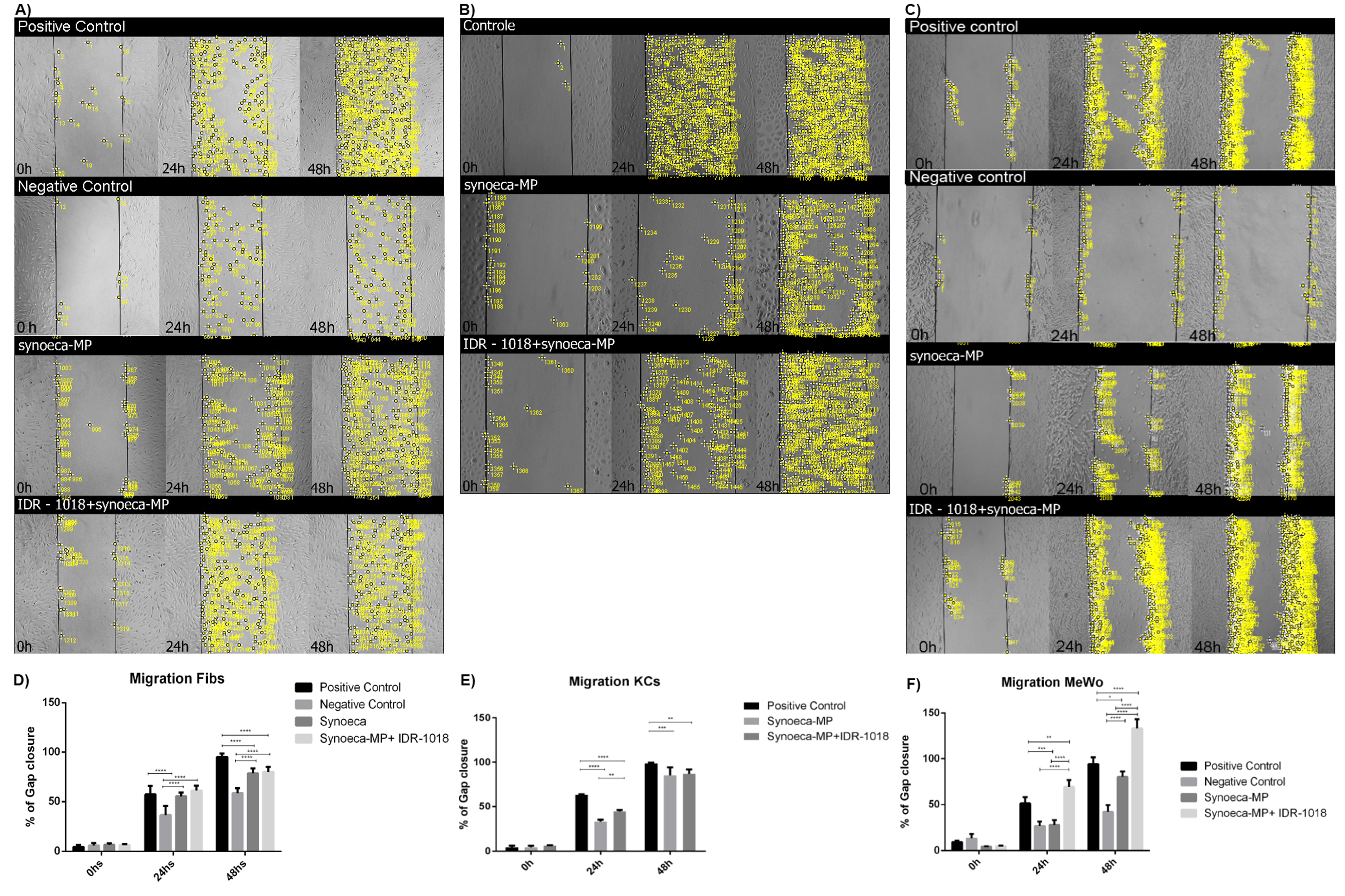
3.4. Effects of Synoeca-MP/IDR-1018 in Cell Proliferation and Migration
3.5. Synoeca-MP/IDR-1018 Can Modify the Expression of Pro-Regenerative Genes
3.6. Synoeca-MP/IDR-1018 Promoted Skin Wound Healing in a 3D Skin Equivalent Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hernandez, R. The use of systemic antibiotics in the treatment of chronic wounds. Dermatol. Ther. 2006, 6, 326–337. [Google Scholar] [CrossRef] [PubMed]
- Maver, T.; Maver, U.; Kleinschek, K.S.; Raščan, I.M.; Smrke, D.M. Advanced therapies of skin injuries. Wien. Klin. Wochenschr. 2015, 127 (Suppl. S5), S187–S198. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.R. Infection in patients with severe burns: Causes and prevention thereof. Infect. Dis. Clin. N. Am. 2007, 3, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Freire, D.; da Cunha, N.; Leite, M.; Kostopoulos, A.; da Silva, S.; de Souza, A.; Nolasco, D.; Franco, O.L.; Mortari, M.; Dias, S.C. Wasp venom peptide, synoeca-MP, from Synoeca surinama shows antimicrobial activity against human and animal pathogenic microorganisms. Pept. Sci. 2020, 112, e24141. [Google Scholar] [CrossRef]
- Dantas, E.M.G.L.; Lima, S.M.F.; Cantuária, A.P.C.; Amorim, I.A.; Almeida, J.A.; Cunha, T.F.; Franco, O.L.; Rezende, T.M.B. Synergistic activity of chlorhexidine and synoeca-MP peptide against Pseudomonas aeruginosa. J. Cell Physiol. 2019, 10, 28265. [Google Scholar] [CrossRef]
- Ormondes de Farias, J.; Resende Ferreira, A.C.; Cardoso Kostopoulos, A.G.; Berto Rezende, T.M.; Dias, S.C. Synergistic activity and immunomodulatory potential of levofloxacin and Synoeca-MP peptide against multi-resistant strains of Klebsiella pneumoniae. Microb. Pathog. 2022, 163, 105403. [Google Scholar] [CrossRef]
- Yanashima, K.; Chieosilapatham, P.; Yoshimoto, E.; Okumura, K.; Ogawa, H.; Niyonsaba, F. Innate defense regulator IDR-1018 activates human mast cells through G protein-, phospholipase C-, MAPK- and NF-ĸB-sensitive pathways. Immunol. Res. 2017, 65, 920–931. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Madera, L.; Afacan, N.; Okumura, K.; Ogawa, H.; Hancock, R.E. The innate defense regulator peptides IDR-HH2, IDR-1002, and IDR-1018 modulate human neutrophil functions. J. Leukoc. Biol. 2013, 94, 159–170. [Google Scholar] [CrossRef]
- Mansour, S.C.; de la Fuente-Núñez, C.; Hancock, R.E. Peptide IDR-1018: Modulating the immune system and targeting bacterial biofilms to treat antibiotic-resistant bacterial infections. J. Pept. Sci. 2015, 21, 323–329. [Google Scholar] [CrossRef]
- Steinstraesser, L.; Hirsch, T.; Schulte, M.; Kueckelhaus, M.; Jacobsen, F.; Mersch, E.A.; Stricker, I.; Afacan, N.; Jenssen, H.; Hancock, R.E.; et al. Innate defense regulator peptide 1018 in wound healing and wound infection. PLoS ONE 2012, 7, e39373. [Google Scholar] [CrossRef]
- Alencar-Silva, T.; Zonari, A.; Foyt, D.; Gang, M.; Pogue, R.; Saldanha-Araujo, F.; Dias, S.C.; Franco, O.L.; Carvalho, J.L. IDR-1018 induces cell proliferation, migration, and reparative gene expression in 2D culture and 3D human skin equivalents. J. Tissue Eng. Regen. Med. 2019, 13, 2018–2030. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.C.; Park, A.Y.; Guan, J.L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Zonari, A.; Brace, L.E.; Al-Katib, K.Z.; Porto, W.F.; Foyt, D.; Guiang, M.; Cruz, E.A.O.; Marshall, B.; Salgueiro, W.G.; Inan, M.D.; et al. Senotherapeutic peptide reduces skin biological age and improves skin health markers. bioRxiv 2020. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Orhan, G.; Bayram, A.; Zer, Y.; Balci, I. Synergy tests by E test and checkerboard methods of antimicrobial combinations against Brucella melitensis. J. Clin. Microbiol. 2005, 43, 140–143. [Google Scholar] [CrossRef]
- Marin-Luevano, P.; Trujillo, V.; Rodriguez-Carlos, A.; González-Curiel, I.; Enciso-Moreno, J.A.; Hancock, R.E.W.; Rivas-Santiago, B. Induction by innate defence regulator peptide 1018 of pro-angiogenic molecules and endothelial cell migration in a high glucose environment. Peptides 2018, 101, 135–144. [Google Scholar] [CrossRef]
- Brook, I. Secondary bacterial infections complicating skin lesions. J. Med. Microbiol. 2002, 51, 808–812. [Google Scholar] [CrossRef]
- Liu, M.; Wang, X.; Li, H.; Xia, C.; Liu, Z.; Liu, J.; Yin, A.; Lou, X.; Wang, H.; Mo, X.; et al. Magnesium oxide-incorporated electrospun membranes inhibit bacterial infections and promote the healing process of infected wounds. J. Mater. Chem. B 2021, 17, 3727–3744. [Google Scholar] [CrossRef]
- Krishna, S.; Miller, L.S. Host-pathogen interactions between the skin and Staphylococcus aureus. Curr. Opin. Microbiol. 2012, 15, 28–35. [Google Scholar] [CrossRef]
- Terreni, M.; Taccani, M.; Pregnolato, M. New Antibiotics for Multidrug-Resistant Bacterial Strains: Latest Research Developments and Future Perspectives. Molecules 2021, 26, 2671. [Google Scholar] [CrossRef]
- Subhan, F.; Kang, H.Y.; Lim, Y.; Ikram, M.; Baek, S.Y.; Jin, S.; Jeong, Y.H.; Kwak, J.Y.; Yoon, S. Fish Scale Collagen Peptides Protect against CoCl2/TNF-α-Induced Cytotoxicity and Inflammation via Inhibition of ROS, MAPK, and NF-κB Pathways in HaCaT Cells. Oxid. Med. Cell Longev. 2017, 2017, 9703609. [Google Scholar] [CrossRef] [PubMed]
- Querobino, S.M.; Costa, M.S.; Alberto-Silva, C. Protective effects of distinct proline-rich oligopeptides from B. jararaca snake venom against oxidative stress-induced neurotoxicity. Toxicon 2019, 167, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Pena, O.M.; Afacan, N.; Pistolic, J.; Chen, C.; Madera, L.; Falsafi, R.; Fjell, C.D.; Hancock, R.E. Synthetic cationic peptide IDR-1018 modulates human macrophage differentiation. PLoS ONE 2013, 8, e52449. [Google Scholar] [CrossRef] [PubMed]
- Rocha, T.; de Souza, B.M.; Palma, M.S.; da Cruz-Höfling, M.A. Myotoxic effects of mastoparan from Polybia paulista (Hymenoptera, Epiponini) wasp venom in mice skeletal muscle. Toxicon 2007, 50, 589–599. [Google Scholar] [CrossRef]
- Pickart, L.; Vasquez-Soltero, J.M.; Margolina, A. The human tripeptide GHK-Cu in prevention of oxidative stress and degenerative conditions of aging: Implications for cognitive health. Oxid. Med. Cell Longev. 2012, 2012, 324832. [Google Scholar] [CrossRef]
- Lu, Y.Y.; Cheng, C.C.; Huang, S.Y.; Chen, Y.C.; Kao, Y.H.; Lin, Y.K.; Higa, S.; Chen, S.A.; Chen, Y.J. Fibroblast Growth Factor 1 Reduces Pulmonary Vein and Atrium Arrhythmogenesis via Modification of Oxidative Stress and Sodium/Calcium Homeostasis. Front. Cardiovasc. Med. 2022, 18, 813589. [Google Scholar] [CrossRef]
- Shen, X.H.; Xu, S.J.; Jin, C.Y.; Ding, F.; Zhou, Y.C.; Fu, G.S. Interleukin-8 prevents oxidative stress-induced human endothelial cell senescence via telomerase activation. Int. Immunopharmacol. 2013, 16, 261–267. [Google Scholar] [CrossRef]
- Abd El-Wahed, A.; Yosri, N.; Sakr, H.H.; Du, M.; Algethami, A.F.M.; Zhao, C.; Abdelazeem, A.H.; Tahir, H.E.; Masry, S.H.D.; Abdel-Daim, M.M.; et al. Wasp Venom Biochemical Components and Their Potential in Biological Applications and Nanotechnological Interventions. Toxins 2021, 12, 206. [Google Scholar] [CrossRef]
- Mu, L.; Tang, J.; Liu, H.; Shen, C.; Rong, M.; Zhang, Z.; Lai, R. A potential wound-healing-promoting peptide from salamander skin. FASEB J. 2014, 28, 3919–3929. [Google Scholar] [CrossRef]
- Song, Y.; Wu, C.; Zhang, X.; Bian, W.; Liu, N.; Yin, S.; Yang, M.; Luo, M.; Tang, J.; Yang, X. A short peptide potentially promotes the healing of skin wound. Biosci. Rep. 2019, 39, BSR20181734. [Google Scholar] [CrossRef]
- Kim, K.K.; Sheppard, D.; Chapman, H.A. TGF-β1 Signaling and Tissue Fibrosis. Cold Spring Harb. Perspect. Biol. 2018, 10, a022293. [Google Scholar] [CrossRef] [PubMed]
- Liarte, S.; Bernabé-García, Á.; Nicolás, F.J. Role of TGF-β in Skin Chronic Wounds: A Keratinocyte Perspective. Cells 2020, 28, 306. [Google Scholar] [CrossRef] [PubMed]
- Diller, R.B.; Tabor, A.J. The Role of the Extracellular Matrix (ECM) in Wound Healing: A Review. Biomimetics 2022, 7, 87. [Google Scholar] [CrossRef] [PubMed]
- Oono, T.; Shirafuji, Y.; Huh, W.K.; Akiyama, H.; Iwatsuki, K. Effects of human neutrophil peptide-1 on the expression of interstitial collagenase and type I collagen in human dermal fibroblasts. Arch. Dermatol. Res. 2002, 294, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, G.; Liu, Y.; Ji, S.; Li, Y.; Xiang, J.; Zhou, L.; Gao, H.; Zhang, W.; Sun, X.; et al. Pleiotropic Roles of CXCR4 in Wound Repair and Regeneration. Front. Immunol. 2021, 28, 668758. [Google Scholar] [CrossRef]
- Fischer, A.M.; Mercer, J.C.; Iyer, A.; Ragin, M.J.; August, A. Regulation of CXC chemokine receptor 4-mediated migration by the Tec family tyrosine kinase ITK. J. Biol. Chem. 2004, 279, 29816–29820. [Google Scholar] [CrossRef]
- Liepelt, A.; Tacke, F. Stromal cell-derived factor-1 (SDF-1) as a target in liver diseases. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G203–G209. [Google Scholar] [CrossRef]
- Guo, R.; Chai, L.; Chen, L.; Chen, W.; Ge, L.; Li, X.; Li, H.; Li, S.; Cao, C. Stromal cell-derived factor 1 (SDF-1) accelerated skin wound healing by promoting the migration and proliferation of epidermal stem cells. In Vitro Cell Dev. Biol. Anim. 2015, 51, 578–585. [Google Scholar] [CrossRef]
- Haensel, D.; Jin, S.; Sun, P.; Cinco, R.; Dragan, M.; Nguyen, Q.; Cang, Z.; Gong, Y.; Vu, R.; MacLean, A.L.; et al. Defining Epidermal Basal Cell States during Skin Homeostasis and Wound Healing Using Single-Cell Transcriptomics. Cell Rep. 2020, 30, 3932–3947.e6. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Agent | MIC (μg/mL) | MBC (μg/mL) |
|---|---|---|
| Synoeca-MP | 16 | 16 |
| IDR-1018 | Nd | Nd |
| Isolated MIC | Tested Concentrations (μg/mL) | |||||
|---|---|---|---|---|---|---|
| Synoeca-MP (16 μg/mL) | 16 | 16 | 16 | 16 | 16 | 16 |
| IDR-1018 (>128 μg/mL) | 2 | 4 | 8 | 16 | 32 | 64 |
| FICI | 0.28 | 0.31 | 0.375 | 0.5 | 0.75 | 1.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alencar-Silva, T.; Díaz-Martín, R.D.; Zonari, A.; Foyt, D.; Guiang, M.; Pogue, R.; Saldanha-Araujo, F.; Dias, S.C.; Franco, O.L.; Carvalho, J.L. The Combination of Synoeca-MP Antimicrobial Peptide with IDR-1018 Stimulates Proliferation, Migration, and the Expression of Pro-Regenerative Genes in Both Human Skin Cell Cultures and 3D Skin Equivalents. Biomolecules 2023, 13, 804. https://doi.org/10.3390/biom13050804
Alencar-Silva T, Díaz-Martín RD, Zonari A, Foyt D, Guiang M, Pogue R, Saldanha-Araujo F, Dias SC, Franco OL, Carvalho JL. The Combination of Synoeca-MP Antimicrobial Peptide with IDR-1018 Stimulates Proliferation, Migration, and the Expression of Pro-Regenerative Genes in Both Human Skin Cell Cultures and 3D Skin Equivalents. Biomolecules. 2023; 13(5):804. https://doi.org/10.3390/biom13050804
Chicago/Turabian StyleAlencar-Silva, Thuany, Rubén D. Díaz-Martín, Alessandra Zonari, Daniel Foyt, Mylieneth Guiang, Robert Pogue, Felipe Saldanha-Araujo, Simoni Campos Dias, Octavio Luiz Franco, and Juliana Lott Carvalho. 2023. "The Combination of Synoeca-MP Antimicrobial Peptide with IDR-1018 Stimulates Proliferation, Migration, and the Expression of Pro-Regenerative Genes in Both Human Skin Cell Cultures and 3D Skin Equivalents" Biomolecules 13, no. 5: 804. https://doi.org/10.3390/biom13050804