
Comparative Anticancer Potential of Green Tea Extract and Epigallocatechin-3-gallate on Breast Cancer Spheroids
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Green Tea Extract
2.3. Characterization of Green Tea Extract Polyphenols by HPLC
2.4. The 2D and 3D Cell Culture
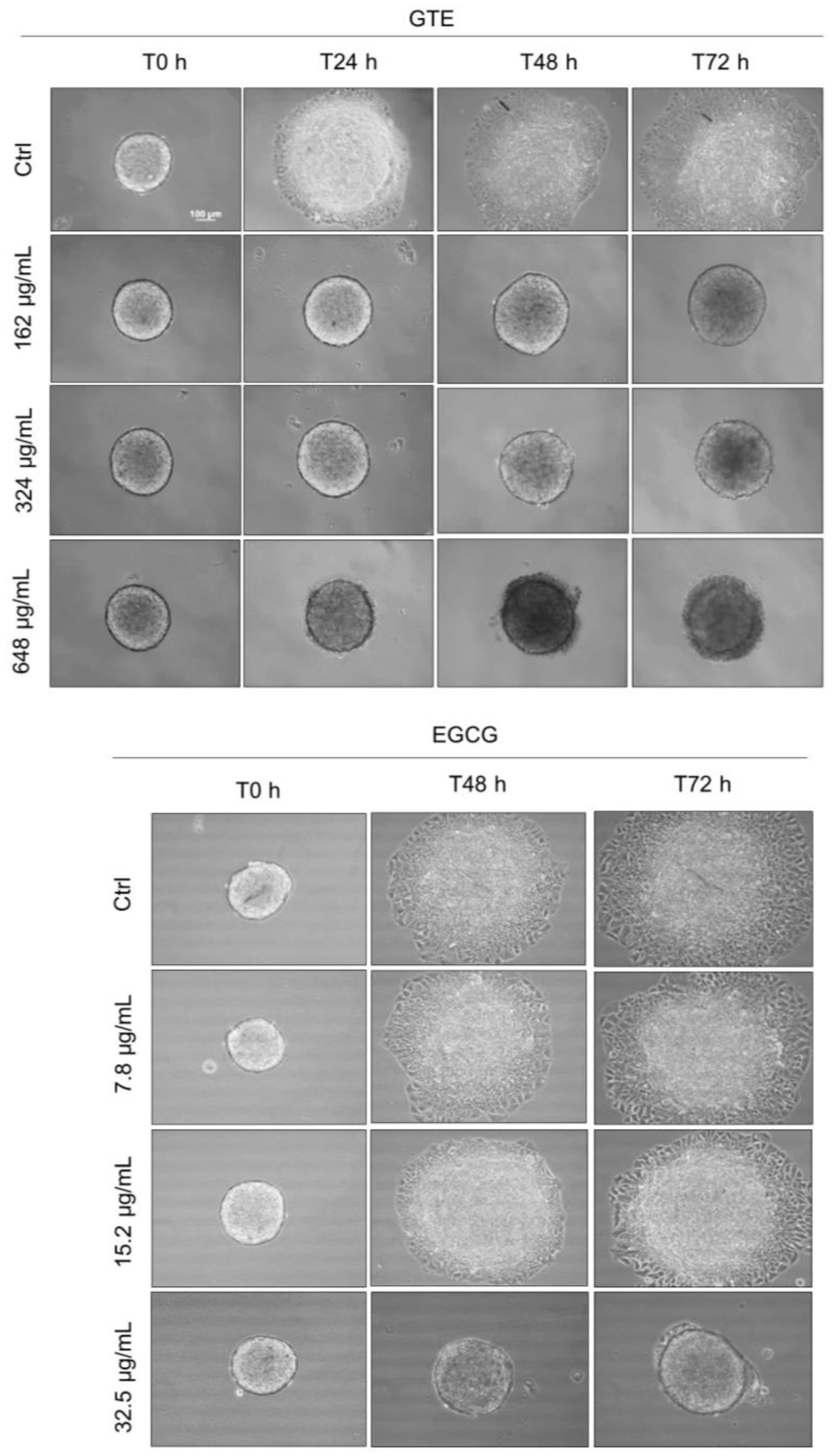
2.5. Spheroid Growth Pattern Follow Up
2.6. Treatment of Spheroids with GTE or EGCG
2.7. Cell Viability Assay
2.8. Migration Assay
2.9. Statistical Analyses
3. Results
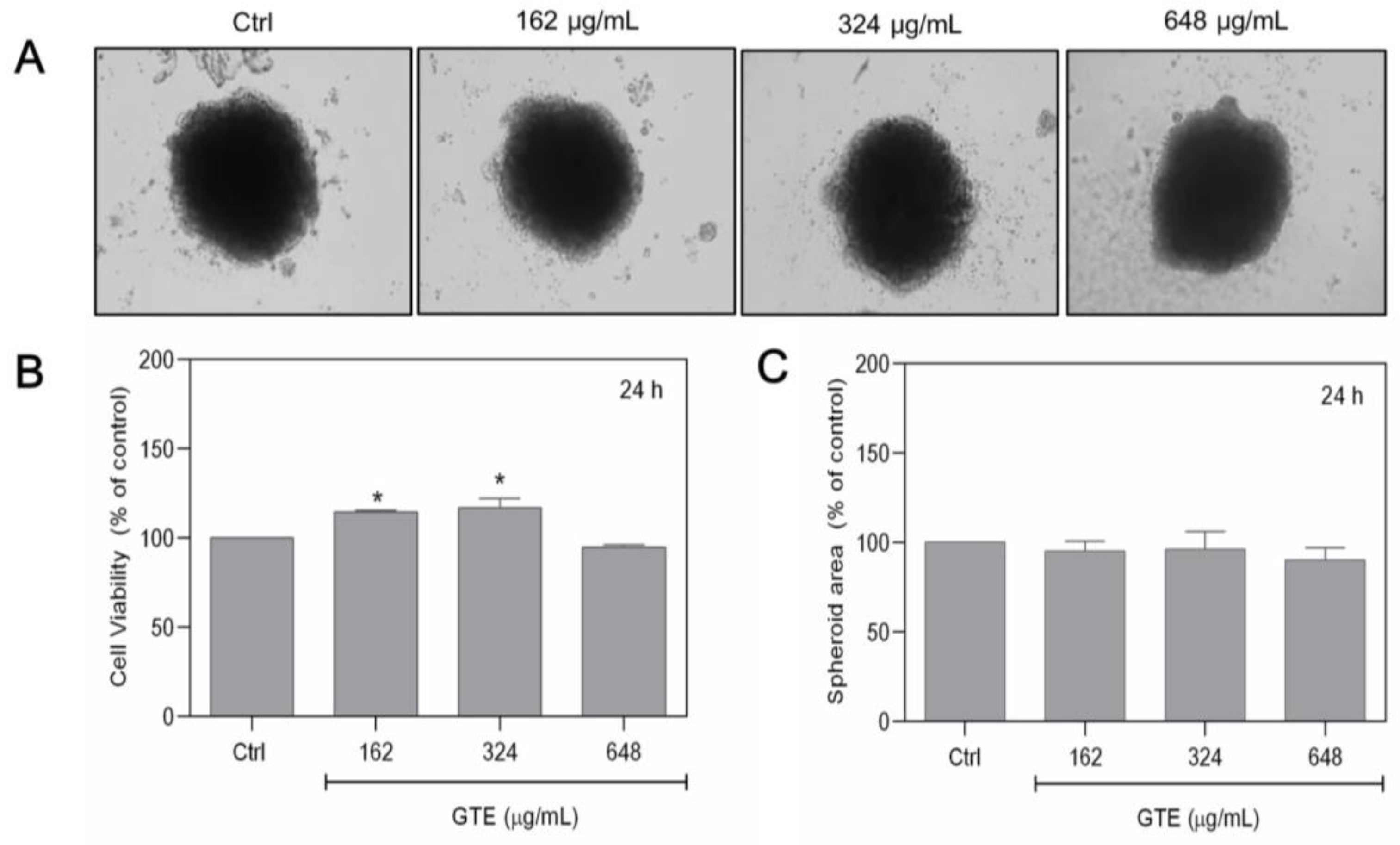
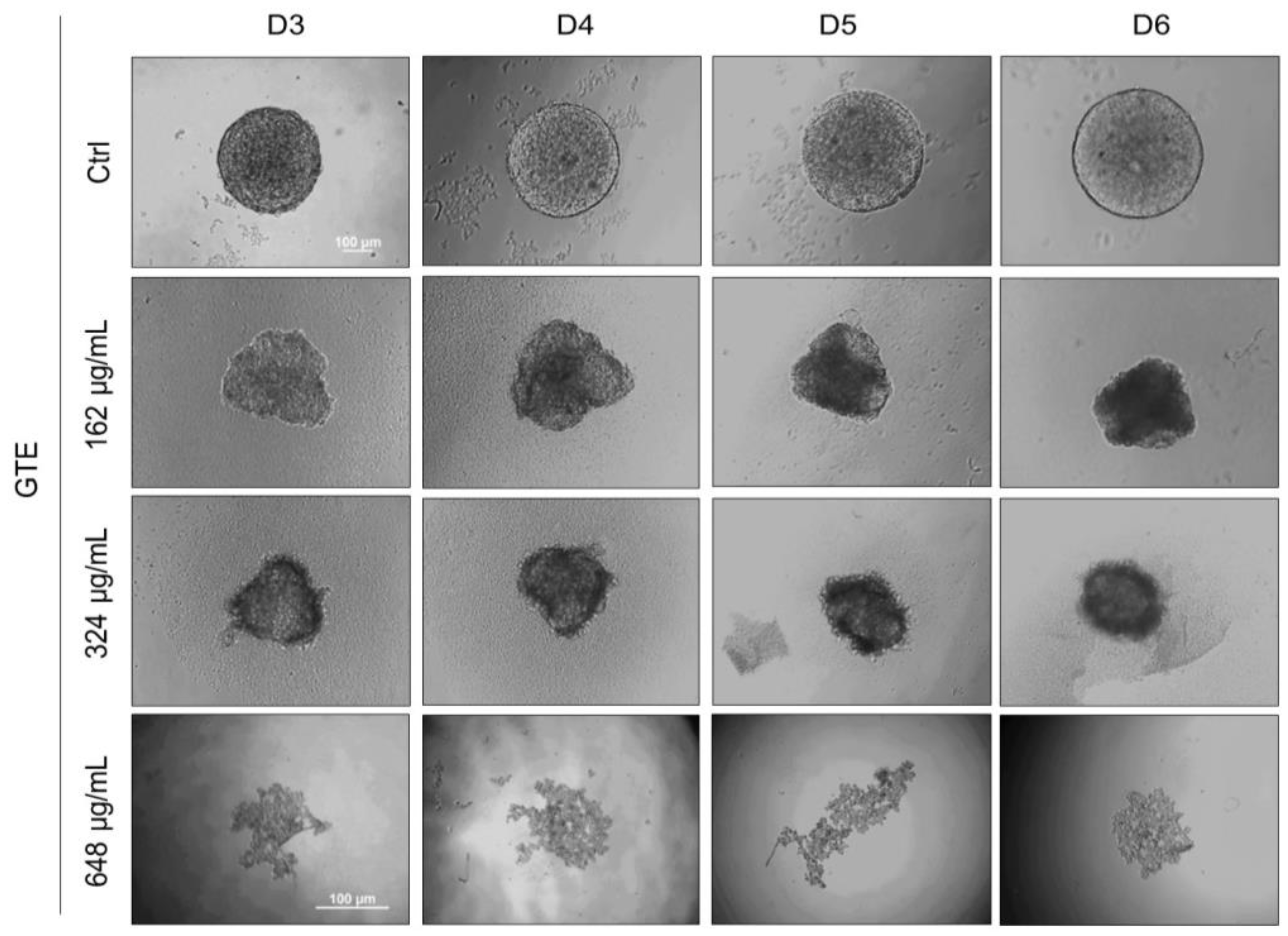
3.1. GTE Affects the Morphology and Growth Pattern of Breast Cancer Spheroids
3.2. GTE Had No Cytotoxic Effects on MCF-10A Cells
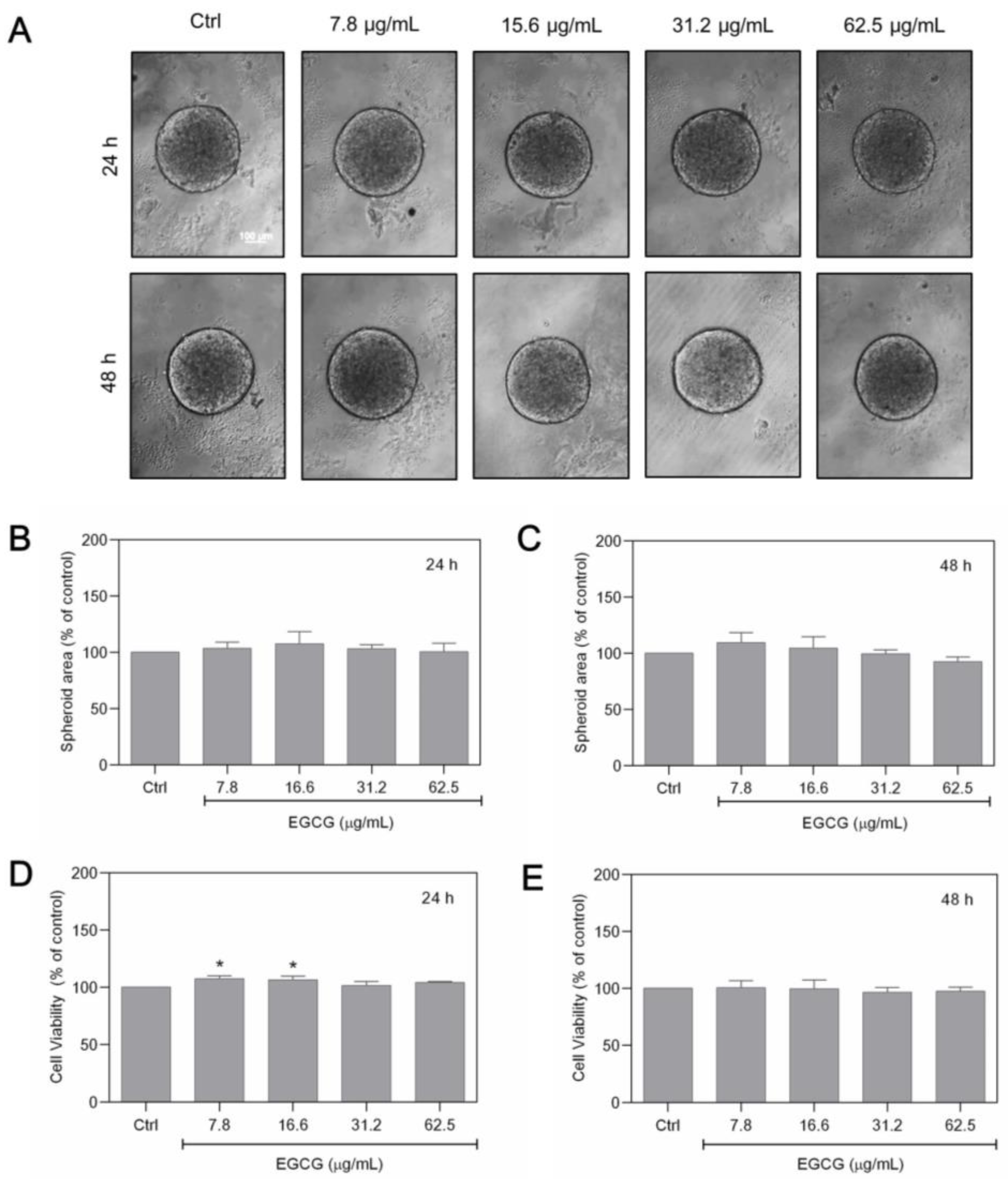
3.3. ECGC Treatment Does Not Promote the Same Effects as GTE
3.4. GTE Inhibits Spheroids’ Formation
3.5. GTE, but Not EGCG, Reduces Spheroids’ Cell Migration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Bag, S.; Mondal, A.; Majumder, A.; Banik, A. Tea and Its Phytochemicals: Hidden Health Benefits & Modulation of Signaling Cascade by Phytochemicals. Food Chem. 2022, 371, 131098. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Shirakami, Y.; Shimizu, M. Possible Mechanisms of Green Tea and Its Constituents against Cancer. Molecules 2018, 23, 2284. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ho, C.T.; Zhou, J.; Santos, J.S.; Armstrong, L.; Granato, D. Chemistry and Biological Activities of Processed Camellia Sinensis Teas: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1474–1495. [Google Scholar] [CrossRef]
- Wang, H.; Helliwell, K.; You, X. Isocratic Elution System for the Determination of Catechins, Caffeine and Gallic Acid in Green Tea Using HPLC. Food Chem. 2000, 68, 115–121. [Google Scholar] [CrossRef]
- Graham, H.N. Green Tea Composition, Consumption and Polyphenol Chemistry. Prev. Med. 1992, 21, 334–350. [Google Scholar] [CrossRef]
- Mocanu, M.M.; Nagy, P.; Szöllosi, J.; Vanden Eynde, J.J.; Mayence, A. Chemoprevention of Breast Cancer by Dietary Polyphenols. Molecules 2015, 20, 22578–22620. [Google Scholar] [CrossRef]
- Cheng, Z.; Zhang, Z.; Han, Y.; Wang, J.; Wang, Y.; Chen, X.; Shao, Y.; Cheng, Y.; Zhou, W.; Lu, X.; et al. A Review on Anti-Cancer Effect of Green Tea Catechins. J. Funct. Foods 2020, 74, 104172. [Google Scholar] [CrossRef]
- Aguilera, J.M. The Food Matrix: Implications in Processing, Nutrition and Health. Crit. Rev. Food Sci. Nutr. 2019, 59, 3612–3629. [Google Scholar] [CrossRef]
- Parada, J.; Aguilera, J.M. Food Microstructure Affects the Bioavailability of Several Nutrients. J. Food Sci. 2007, 72, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Melse-Boonstra, A. Bioavailability of Micronutrients from Nutrient-Dense Whole Foods: Zooming in on Dairy, Vegetables, and Fruits. Front. Nutr. 2020, 7, 101. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.; Amaro, A.L.; Pintado, M. Impact of Food Matrix Components on Nutritional and Functional Properties of Fruit-Based Products. Curr. Opin. Food Sci. 2018, 22, 153–159. [Google Scholar] [CrossRef]
- Nakagawa, K.; Nakayama, K.; Nakamura, M.; Sookwong, P.; Tsuduki, T.; Niino, H.; Kimura, F.; Miyazawa, T. Effects of Co-Administration of Tea Epigallocatechin-3-Gallate (EGCG) and Caffeine on Absorption and Metabolism of EGCG in Humans. Biosci. Biotechnol. Biochem. 2009, 73, 2014–2017. [Google Scholar] [CrossRef]
- Jacobs, D.R.; Tapsell, L.C. Food, Not Nutrients, Is the Fundamental Unit in Nutrition. Nutr. Rev. 2007, 65, 439–450. [Google Scholar] [CrossRef]
- Kapinova, A.; Stefanicka, P.; Kubatka, P.; Zubor, P.; Uramova, S.; Kello, M.; Mojzis, J.; Blahutova, D.; Qaradakhi, T.; Zulli, A.; et al. Are Plant-Based Functional Foods Better Choice against Cancer than Single Phytochemicals? A Critical Review of Current Breast Cancer Research. Biomed. Pharmacother. 2017, 96, 1465–1477. [Google Scholar] [CrossRef]
- Krishnan, S.P. Breast Cancer Heterogeneity: A Focus on Epigenetics and in Vitro 3D Model Systems. Cell J. 2018, 20, 302. [Google Scholar] [CrossRef]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Bliźniak, R.; Łuczewski, Ł.; Lamperska, K. 2D and 3D Cell Cultures—A Comparison of Different Types of Cancer Cell Cultures. Arch. Med. Sci. 2018, 14, 910–919. [Google Scholar] [CrossRef]
- Millard, M.; Yakavets, I.; Zorin, V.; Kulmukhamedova, A.; Marchal, S.; Bezdetnaya, L. Drug Delivery to Solid Tumors: The Predictive Value of the Multicellular Tumor Spheroid Model for Nanomedicine Screening. Int. J. Nanomed. 2017, 12, 7993–8007. [Google Scholar] [CrossRef]
- Imamura, Y.; Mukohara, T.; Shimono, Y.; Funakoshi, Y.; Chayahara, N.; Toyoda, M.; Kiyota, N.; Takao, S.; Kono, S.; Nakatsura, T.; et al. Comparison of 2D- and 3D-Culture Models as Drug-Testing Platforms in Breast Cancer. Oncol. Rep. 2015, 33, 1837–1843. [Google Scholar] [CrossRef]
- Nishiyama, M.F.; Costa, M.A.F.; da Costa, A.M.; de Souza, C.G.M.; Bôer, C.G.; Bracht, C.K.; Peralta, R.M. Chá Verde Brasileiro (Camellia sinensis Var Assamica): Efeitos Do Tempo de Infusão, Acondicionamento Da Erva e Forma de Preparo Sobre a Eficiência de Extração Dos Bioativos e Sobre a Estabilidade Da Bebida. Food Sci. Technol. 2010, 30, 191–196. [Google Scholar] [CrossRef]
- Friedrich, J.; Seidel, C.; Ebner, R.; Kunz-Schughart, L.A. Spheroid-Based Drug Screen: Considerations and Practical Approach. Nat. Protoc. 2009, 4, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Eilenberger, C.; Kratz, S.R.A.; Rothbauer, M.; Ehmoser, E.K.; Ertl, P.; Küpcü, S. Optimized AlamarBlue Assay Protocol for Drug Dose-Response Determination of 3D Tumor Spheroids. MethodsX 2018, 5, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Vinci, M.; Box, C.; Zimmermann, M.; Eccles, S.A. Tumor Spheroid-Based Migration Assays for Evaluation of Therapeutic Agents. Target. Identif. Valid. Drug Discov. Methods Protoc. 2013, 253–266. [Google Scholar] [CrossRef]
- Hussain, S.S.; Kumar, A.P.; Ghosh, R. Food-Based Natural Products for Cancer Management: Is the Whole Greater than the Sum of the Parts? Semin. Cancer Biol. 2016, 40, 233–246. [Google Scholar] [CrossRef] [PubMed]
- Tungmunnithum, D.; Thongboonyou, A.; Pholboon, A.; Yangsabai, A. Flavonoids and Other Phenolic Compounds from Medicinal Plants for Pharmaceutical and Medical Aspects: An Overview. Medicines 2018, 5, 93. [Google Scholar] [CrossRef]
- Yu, S.; Zhu, L.; Wang, K.; Yan, Y.; He, J.; Ren, Y. Green Tea Consumption and Risk of Breast Cancer: A Systematic Review and Updated Meta-Analysis of Case-Control Studies. Medicine 2019, 98, e16147. [Google Scholar] [CrossRef]
- Xing, L.; Zhang, H.; Qi, R.; Tsao, R.; Mine, Y. Recent Advances in the Understanding of the Health Benefits and Molecular Mechanisms Associated with Green Tea Polyphenols. J. Agric. Food Chem. 2019, 67, 1029–1043. [Google Scholar] [CrossRef]
- Crespy, V.; Williamson, G. A Review of the Health Effects of Green Tea Catechins in In Vivo Animal Models. J. Nutr. 2018, 134, 3431S–3440S. [Google Scholar] [CrossRef]
- Huang, C.Y.; Han, Z.; Li, X.; Xie, H.H.; Zhu, S.S. Mechanism of Egcg Promoting Apoptosis of MCF–7 Cell Line in Human Breast Cancer. Oncol. Lett. 2017, 14, 3623–3627. [Google Scholar] [CrossRef]
- Moradzadeh, M.; Hosseini, A.; Erfanian, S.; Rezaei, H. Epigallocatechin-3-Gallate Promotes Apoptosis in Human Breast Cancer T47D Cells through down-Regulation of PI3K/AKT and Telomerase. Pharmacol. Rep. 2017, 69, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Knaze, V.; Zamora-Ros, R. Polyphenols: Dietary Assessment and Role in the Prevention of Cancers. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 512–521. [Google Scholar] [CrossRef] [PubMed]
- Samavat, H.; Ursin, G.; Emory, T.H.; Lee, E.; Wang, R.; Torkelson, C.J.; Dostal, A.M.; Swenson, K.; Le, C.T.; Yang, C.S.; et al. A Randomized Controlled Trial of Green Tea Extract Supplementation and Mammographic Density in Postmenopausal Women at Increased Risk of Breast Cancer. Cancer Prev. Res. 2017, 10, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Ifie, I.; Marshall, L.J. Food Processing and Its Impact on Phenolic Constituents in Food. Cogent Food Agric. 2018, 4, 1507782. [Google Scholar] [CrossRef]
- Harrisson, R.G. Observations on the Living Developing Nerve Fiber. Soc. Exp. Biol. Med. 1906, 98, 140–143. [Google Scholar] [CrossRef]
- Hoarau-Véchot, J.; Rafii, A.; Touboul, C.; Pasquier, J. Halfway between 2D and Animal Models: Are 3D Cultures the Ideal Tool to Study Cancer-Microenvironment Interactions? Int. J. Mol. Sci. 2018, 19, 181. [Google Scholar] [CrossRef] [PubMed]
- Sachinidis, A.; Seul, C.; Seewald, S.; Ahn, H.Y.; Ko, Y.; Vetter, H. Green Tea Compounds Inhibit Tyrosine Phosphorylation of PDGF β-Receptor and Transformation of A172 Human Glioblastoma. FEBS Lett. 2000, 471, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Do Amaral, J.B.; Rezende-Teixeira, P.; Freitas, V.M.; MacHado-Santelli, G.M. MCF-7 Cells as a Three-Dimensional Model for the Study of Human Breast Cancer. Tissue Eng. Part C Methods 2011, 17, 1097–1107. [Google Scholar] [CrossRef]
- Kenny, P.A.; Lee, G.Y.; Myers, C.A.; Neve, R.M.; Semeiks, J.R.; Spellman, P.T.; Lorenz, K.; Lee, E.H.; Barcellos-Hoff, M.H.; Petersen, O.W.; et al. The Morphologies of Breast Cancer Cell Lines in Three-Dimensional Assays Correlate with Their Profiles of Gene Expression. Mol. Oncol. 2007, 1, 84–96. [Google Scholar] [CrossRef]
- Santos, R.A.; Andrade, E.D.S.; Monteiro, M.; Fialho, E.; Silva, J.L.; Daleprane, J.B.; da Costa, D.C.F. Green Tea (Camellia sinensis) Extract Induces P53-Mediated Cytotoxicity and Inhibits Migration of Breast Cancer Cells. Foods 2021, 10, 3154. [Google Scholar] [CrossRef]
- Mueller-Klieser, W.; Schreiber-Klais, S.; Walenta, S.; Kreuter, M.H. Bioactivity of Well-Defined Green Tea Extracts in Multicellular Tumor Spheroids. Int. J. Oncol. 2002, 21, 1307–1315. [Google Scholar] [CrossRef] [PubMed]
- Günther, S.; Ruhe, C.; Derikito, M.G.; Böse, G.; Sauer, H.; Wartenberg, M. Polyphenols Prevent Cell Shedding from Mouse Mammary Cancer Spheroids and Inhibit Cancer Cell Invasion in Confrontation Cultures Derived from Embryonic Stem Cells. Cancer Lett. 2007, 250, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Safari, F.; Azad, N.R.; Ezdiny, A.A.; Pakizehkar, S.; Koohpar, Z.K.; Ranji, N. Antitumor Activities of Green Tea by Up-Regulation of MiR-181a Expression in LNCaP Cells Using 3D Cell Culture Model. Avicenna J. Med. Biotechnol. 2021, 14, 89. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Isemura, M. Inhibitory Effect of Epigallocatechin Gallate on Adhesion of Murine Melanoma Cells to Laminin. Cancer Lett. 2001, 173, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Lo, H.M.; Hung, C.F.; Huang, Y.Y.; Wu, W.B. Tea Polyphenols Inhibit Rat Vascular Smooth Muscle Cell Adhesion and Migration on Collagen and Laminin via Interference with Cell-ECM Interaction. J. Biomed. Sci. 2007, 14, 637–645. [Google Scholar] [CrossRef]
- Li, Y.; Kuscu, C.; Banach, A.; Zhang, Q.; Pulkoski-Gross, A.; Kim, D.; Liu, J.; Roth, E.; Li, E.; Shroyer, K.R.; et al. MicroRNA-181a-5p Inhibits Cancer Cell Migration and Angiogenesis via Downregulation of Matrix Metalloproteinase-14. Cancer Res. 2016, 75, 2674–2685. [Google Scholar] [CrossRef]
- Seo, E.J.; Wu, C.F.; Ali, Z.; Wang, Y.H.; Khan, S.I.; Walker, L.A.; Khan, I.A.; Efferth, T. Both Phenolic and Non-Phenolic Green Tea Fractions Inhibit Migration of Cancer Cells. Front. Pharmacol. 2016, 7, 398. [Google Scholar] [CrossRef]
- Luo, K.W.; Xia, J.; Cheng, B.H.; Gao, H.C.; Fu, L.W.; Luo, X. Le Tea Polyphenol EGCG Inhibited Colorectal-Cancer-Cell Proliferation and Migration via Downregulation of STAT3. Gastroenterol. Rep. 2021, 9, 59–70. [Google Scholar] [CrossRef]
- Panji, M.; Behmard, V.; Zare, Z.; Malekpour, M.; Nejadbiglari, H.; Yavari, S.; Nayerpour dizaj, T.; Safaeian, A.; Maleki, N.; Abbasi, M.; et al. Suppressing Effects of Green Tea Extract and Epigallocatechin-3-Gallate (EGCG) on TGF-β- Induced Epithelial-to-Mesenchymal Transition via ROS/Smad Signaling in Human Cervical Cancer Cells. Gene 2021, 794, 145774. [Google Scholar] [CrossRef]
- Deb, G.; Thakur, V.S.; Limaye, A.M.; Gupta, S. Epigenetic Induction of Tissue Inhibitor of Matrix Metalloproteinase-3 by Green Tea Polyphenols in Breast Cancer Cells. Mol. Carcinog. 2015, 54, 485–499. [Google Scholar] [CrossRef]
- Somiari, S.B.; Shriver, C.D.; Heckman, C.; Olsen, C.; Hu, H.; Jordan, R.; Arciero, C.; Russell, S.; Garguilo, G.; Hooke, J.; et al. Plasma Concentration and Activity of Matrix Metalloproteinase 2 and 9 in Patients with Breast Disease, Breast Cancer and at Risk of Developing Breast Cancer. Cancer Lett. 2006, 233, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Morin, M.P.; Grenier, D. Regulation of Matrix Metalloproteinase Secretion by Green Tea Catechins in a Three-Dimensional Co-Culture Model of Macrophages and Gingival Fibroblasts. Arch. Oral. Biol. 2017, 75, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Spallarossa, P.; Altieri, P.; Garibaldi, S.; Ghigliotti, G.; Barisione, C.; Manca, V.; Fabbi, P.; Ballestrero, A.; Brunelli, C.; Barsotti, A. Matrix Metalloproteinase-2 and -9 Are Induced Differently by Doxorubicin in H9c2 Cells: The Role of MAP Kinases and NAD(P)H Oxidase. Cardiovasc. Res. 2006, 69, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Dusemund, B.; Filipič, M.; Frutos, M.J.; Galtier, P.; Gott, D.; Gundert-Remy, U.; et al. Scientific Opinion on the Safety of Green Tea Catechins. EFSA J. 2018, 16, e05239. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Webster, D.; Cao, J.; Shao, A. The Safety of Green Tea and Green Tea Extracts Consumption in Adults—Results of a Systematic Review. Regul. Toxicol. Pharmacol. 2018, 95, 412–433. [Google Scholar] [CrossRef] [PubMed]
- Breslin, S.; O’Driscoll, L. The Relevance of Using 3D Cell Cultures, in Addition to 2D Monolayer Cultures, When Evaluating Breast Cancer Drug Sensitivity and Resistance. Oncotarget 2016, 7, 45745. [Google Scholar] [CrossRef]
- Zimmermann, M.; Box, C.; Eccles, S.A. Two-Dimensional vs. Three-Dimensional in Vitro Tumor Migration and Invasion Assays. In Methods in Molecular Biology; Springer: Berlin/Heidelberg, Germany, 2013; Volume 986, pp. 227–252. ISBN 9781627033107. [Google Scholar]
- Grantab, R.; Sivananthan, S.; Tannock, I.F. The Penetration of Anticancer Drugs through Tumor Tissue as a Function of Cellular Adhesion and Packing Density of Tumor Cells. Cancer Res. 2006, 66, 1033–1039. [Google Scholar] [CrossRef]
- Tanner, K.; Gottesman, M.M. Beyond 3D Culture Models of Cancer. Sci. Transl. Med. 2015, 7, 283ps9. [Google Scholar] [CrossRef]
- Samanta, S. Potential Bioactive Components and Health Promotional Benefits of Tea (Camellia Sinensis). J. Am. Coll. Nutr. 2020, 41, 65–93. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, R.A.; Pessoa, H.R.; Daleprane, J.B.; de Faria Lopes, G.P.; da Costa, D.C.F. Comparative Anticancer Potential of Green Tea Extract and Epigallocatechin-3-gallate on Breast Cancer Spheroids. Foods 2024, 13, 64. https://doi.org/10.3390/foods13010064
Santos RA, Pessoa HR, Daleprane JB, de Faria Lopes GP, da Costa DCF. Comparative Anticancer Potential of Green Tea Extract and Epigallocatechin-3-gallate on Breast Cancer Spheroids. Foods. 2024; 13(1):64. https://doi.org/10.3390/foods13010064
Chicago/Turabian StyleSantos, Ronimara A., Heloisa Rodrigues Pessoa, Julio Beltrame Daleprane, Giselle Pinto de Faria Lopes, and Danielly C. Ferraz da Costa. 2024. "Comparative Anticancer Potential of Green Tea Extract and Epigallocatechin-3-gallate on Breast Cancer Spheroids" Foods 13, no. 1: 64. https://doi.org/10.3390/foods13010064